

Genetic variation of the IL-28B promoter affecting gene expression
- PMID: 22046316
- PMCID: PMC3201970
- DOI: 10.1371/journal.pone.0026620
Genetic variation of the IL-28B promoter affecting gene expression
Abstract
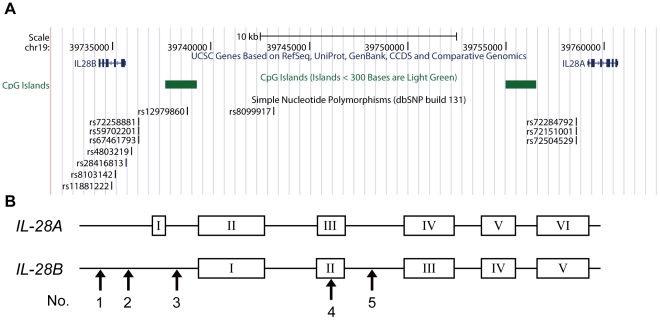
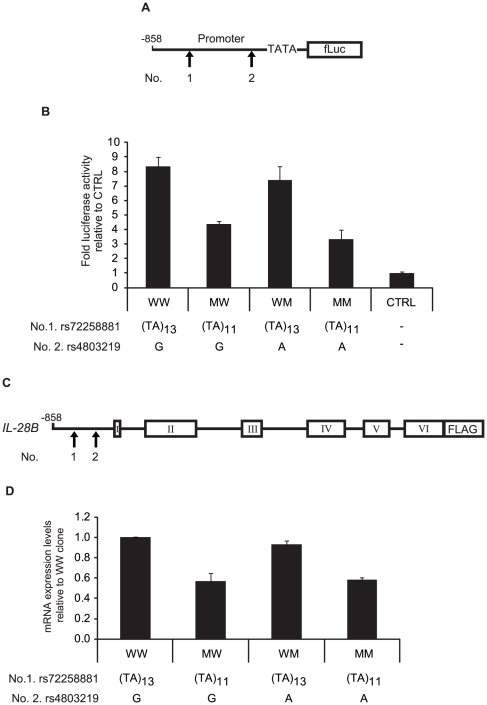
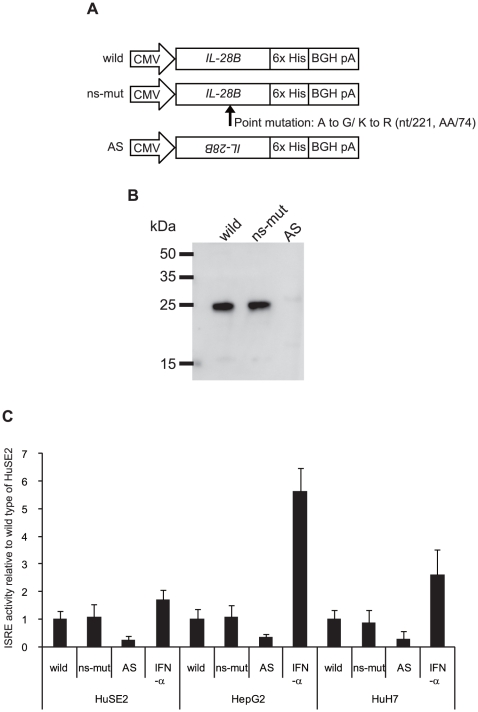
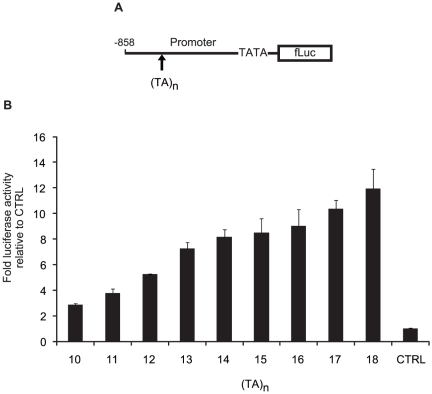
The current standard of care for the treatment of chronic hepatitis C is pegylated interferon-α (PEG-IFNα) and ribavirin (RBV). The treatment achieves a sustained viral clearance in only approximately 50% of patients. Recent whole genome association studies revealed that single nucleotide polymorphisms (SNPs) around IL-28B have been associated with response to the standard therapy and could predict treatment responses at approximately 80%. However, it is not clear which SNP is most informative because the genomic region containing significant SNPs shows strong linkage disequilibrium. We focused on SNPs in close proximity to the IL-28B gene to evaluate the function of each and identify the SNP affecting the IL-28B expression level most. The structures of IL-28A/B from 5' to 3'-UTR were determined by complete cDNA cloning. Both IL-28A and 28B genes consisted of 6 exons, differing from the CCDS data of NCBI. Two intron SNPs and a nonsynonymous SNP did not affect IL-28B gene function and expression levels but a SNP located in the proximal promoter region influenced gene expression. A (TA) dinucleotide repeat, rs72258881, located in the promoter region was discovered by our functional studies of the proximal SNPs upstream of IL-28B; the transcriptional activity of the promoter increased gradually in a (TA)(n) length-dependent manner following IFN-α and lipopolysaccharide stimulation. Healthy Japanese donors exhibited a broad range of (TA) dinucleotide repeat numbers from 10 to 18 and the most prevalent genotype was 12/12 (75%), differing from the database (13/13). However, genetic variation of IL-28A corresponding to that of IL-28B was not detected in these Japanese donors. These findings suggest that the dinucleotide repeat could be associated with the transcriptional activity of IL-28B as well as being a marker to improve the prediction of the response to interferon-based hepatitis C virus treatment.
Conflict of interest statement
Figures

References
-
- Kotenko SV, Gallagher G, Baurin VV, Lewis-Antes A, Shen M, et al. IFN-lambdas mediate antiviral protection through a distinct class II cytokine receptor complex. Nat Immunol. 2003;4:69–77. - PubMed
-
- Sheppard P, Kindsvogel W, Xu W, Henderson K, Schlutsmeyer S, et al. IL-28, IL-29 and their class II cytokine receptor IL-28R. Nat Immunol. 2003;4:63–68. - PubMed
-
- Doyle SE, Schreckhise H, Khuu-Duong K, Henderson K, Rosler R, et al. Interleukin-29 uses a type 1 interferon-like program to promote antiviral responses in human hepatocytes. Hepatology. 2006;44:896–906. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
