

Antimicrobial peptides inhibit polyinosinic-polycytidylic acid-induced immune responses
- PMID: 22048772
- PMCID: PMC3233200
- DOI: 10.4049/jimmunol.1102144
Antimicrobial peptides inhibit polyinosinic-polycytidylic acid-induced immune responses
Abstract
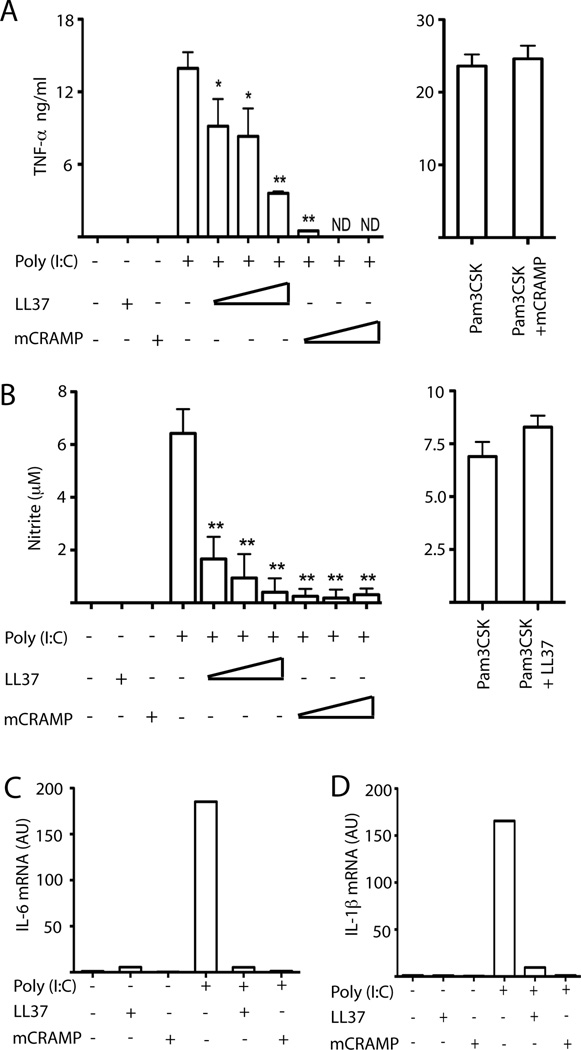
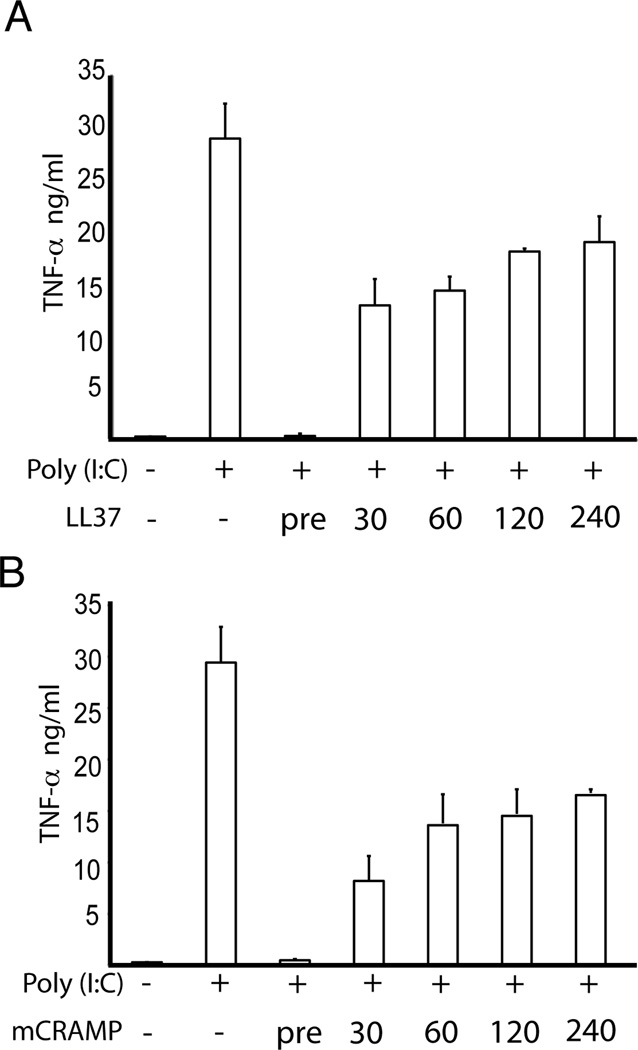
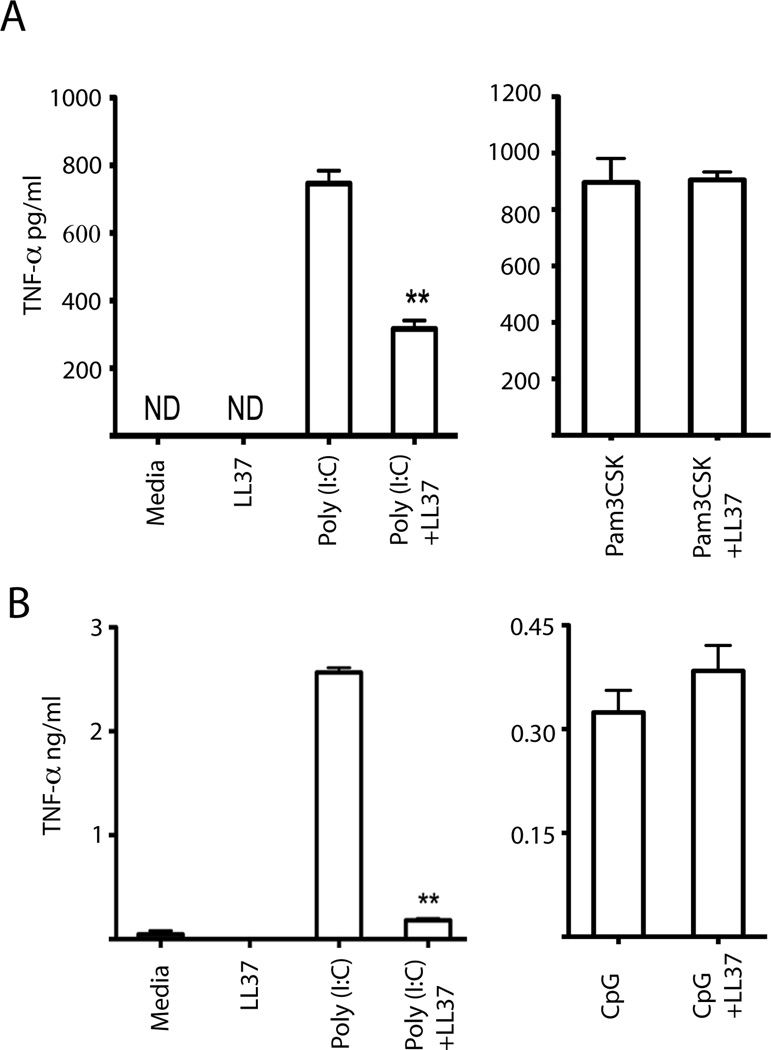
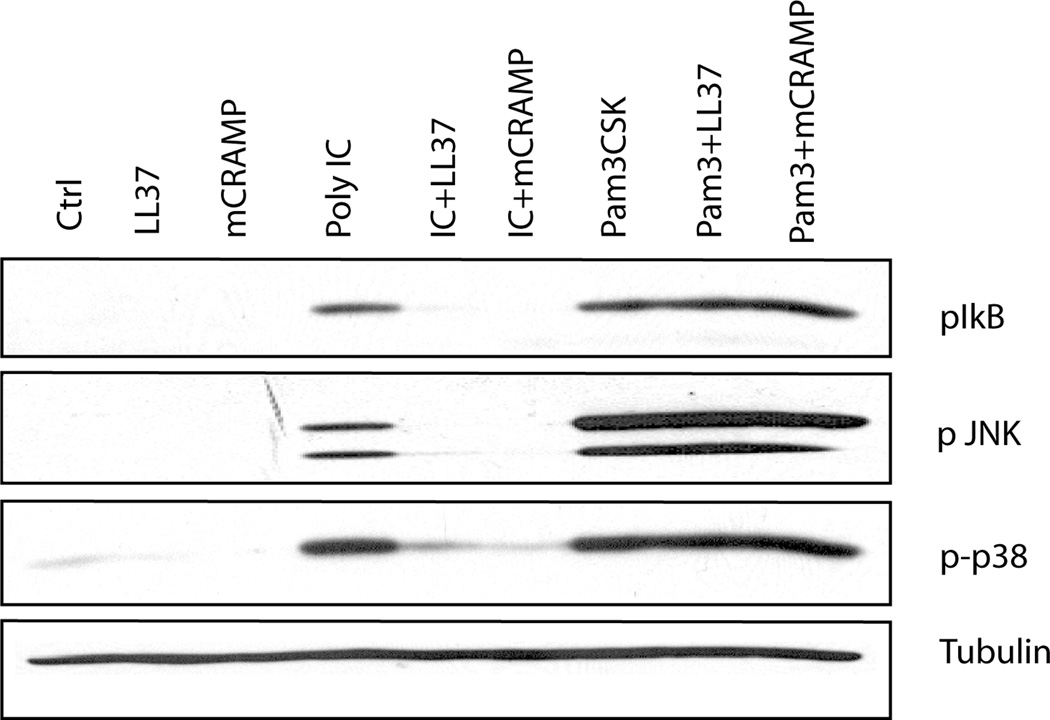
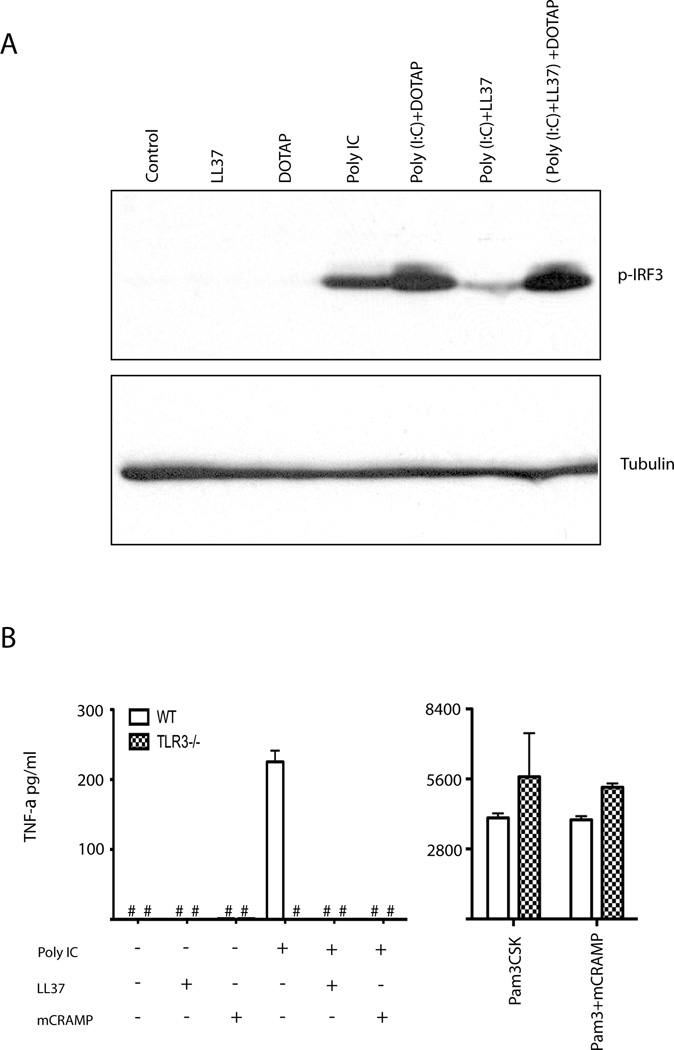
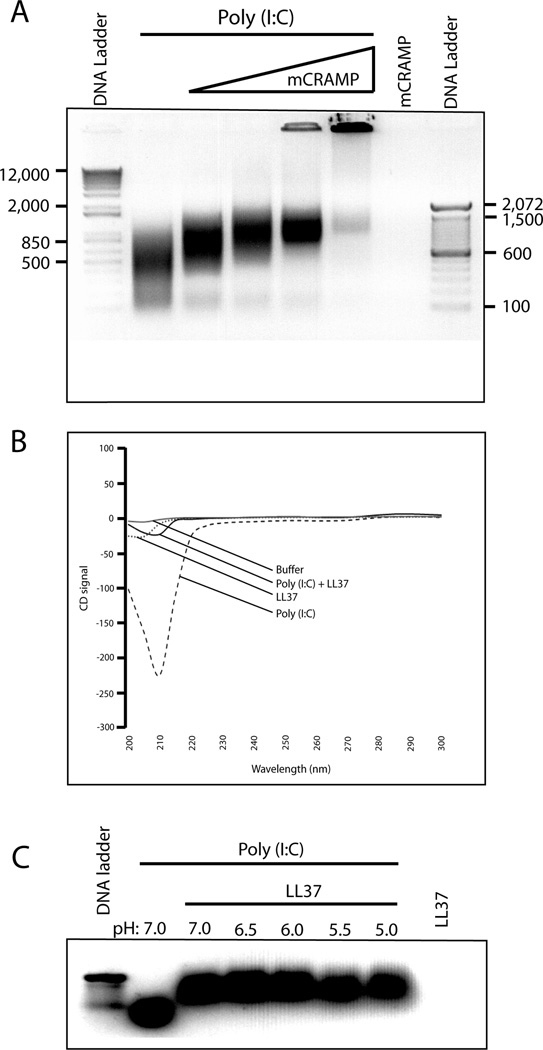
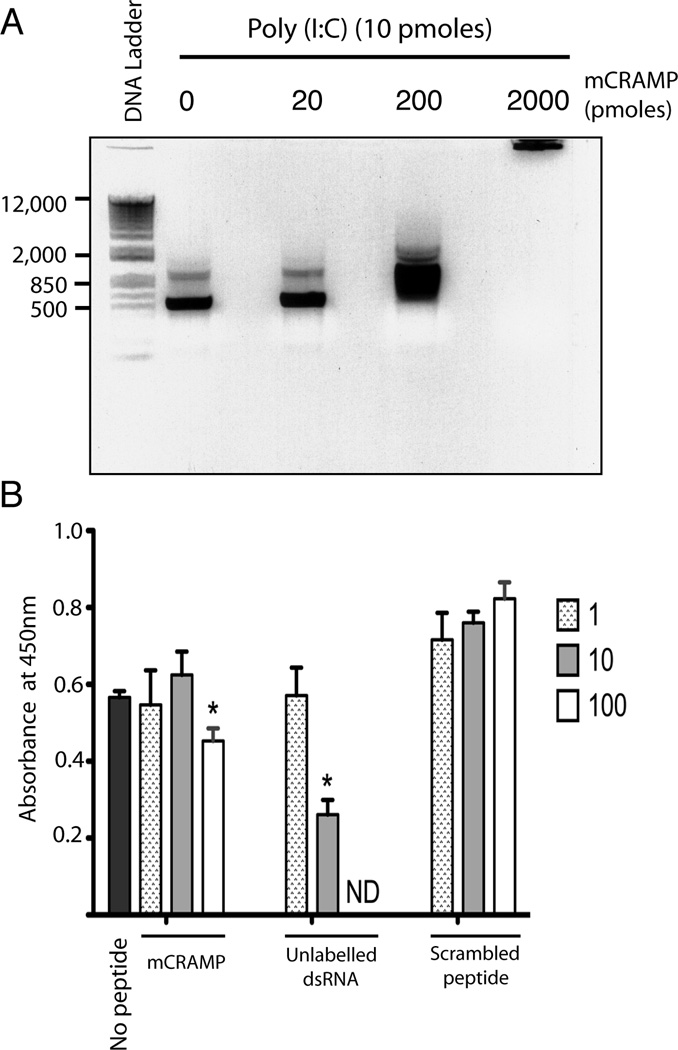
Viral proteins and nucleic acids stimulate TLRs to elicit production of cytokines, chemokines, and IFNs. Because of their immunostimulatory activity, several TLR agonists are being developed as vaccine adjuvants and cancer immunotherapeutics. However, TLR signaling is modified by disease state, which could enhance or impair therapeutic efficacy. For example, in the skin of psoriasis patients, the human cationic antimicrobial peptide LL37 is highly expressed and binds to host DNA. Association with LL37 enhances DNA uptake into intracellular compartments, where it stimulates TLR9-dependent overproduction of IFNs. Polyinosinic-polycytidylic acid (poly(I:C)), an analog of viral dsRNA, is recognized by TLR3 and is currently in preclinical trials as an inducer of type I IFN. If LL37 similarly enhanced IFN production, use of poly(I:C) might be contraindicated in certain conditions where LL37 is elevated. In this study, we show that TLR3 signaling was not enhanced, but was dramatically inhibited, by LL37 or mouse cathelicidin-related antimicrobial peptide in macrophages, microglial cells, and dendritic cells. Inhibition correlated with formation of a strong complex between antimicrobial peptides and poly(I:C), which partially inhibited poly(I:C) binding to TLR3. Therefore, after injury or during existing acute or chronic inflammation, when LL37 levels are elevated, the therapeutic activity of poly(I:C) will be compromised. Our findings highlight the importance of using caution when therapeutically delivering nucleic acids as immunomodulators.
Figures
References
-
- Akira S, Takeda K, Kaisho T. Toll-like receptors: critical proteins linking innate and acquired immunity. Nat Immunol. 2001;2:675–680. - PubMed
-
- O'Neill LA, Fitzgerald KA, Bowie AG. The Toll-IL-1 receptor adaptor family grows to five members. Trends Immunol. 2003;24:286–290. - PubMed
-
- Takeda K, Kaisho T, Akira S. Toll-like receptors. Annu Rev Immunol. 2003;21:335–376. - PubMed
-
- Kim YM, Brinkmann MM, Paquet ME, Ploegh HL. UNC93B1 delivers nucleotide-sensing toll-like receptors to endolysosomes. Nature. 2008;452:234–238. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
