

Dynamic analysis of amyloid β-protein in behaving mice reveals opposing changes in ISF versus parenchymal Aβ during age-related plaque formation
- PMID: 22049429
- PMCID: PMC3227224
- DOI: 10.1523/JNEUROSCI.3272-11.2011
Dynamic analysis of amyloid β-protein in behaving mice reveals opposing changes in ISF versus parenchymal Aβ during age-related plaque formation
Abstract
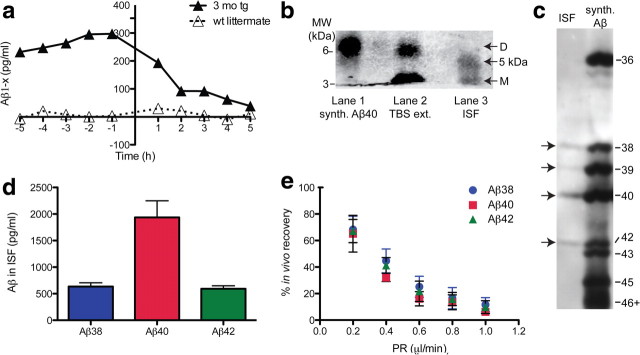
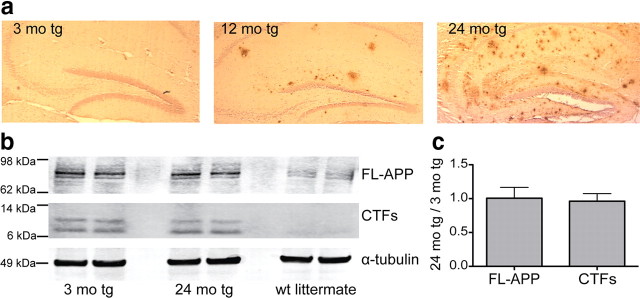
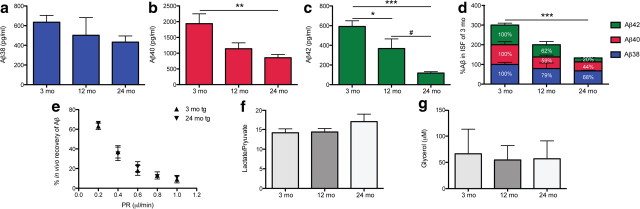
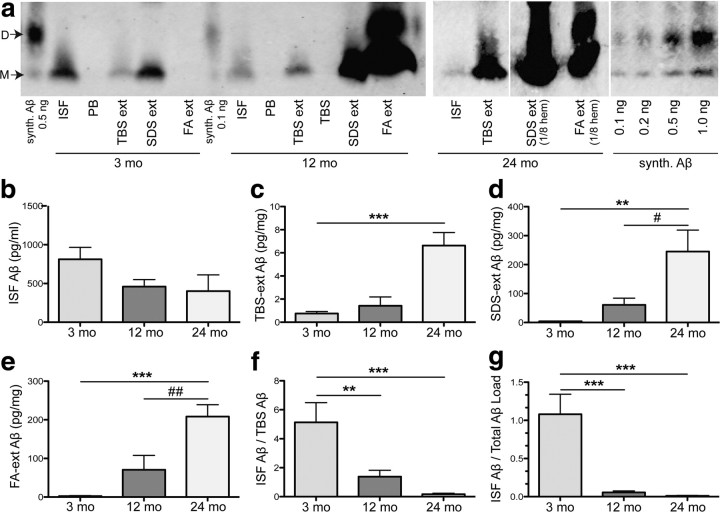
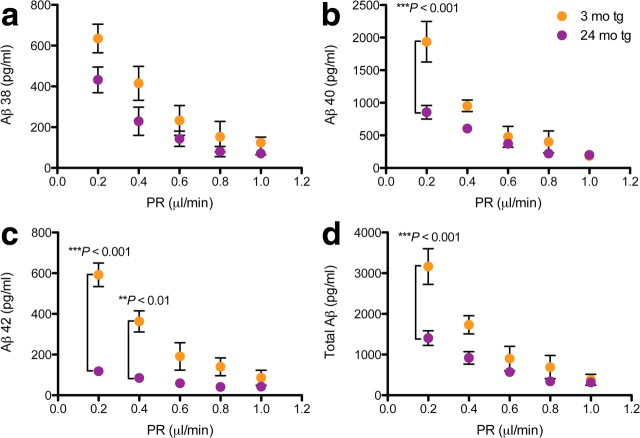
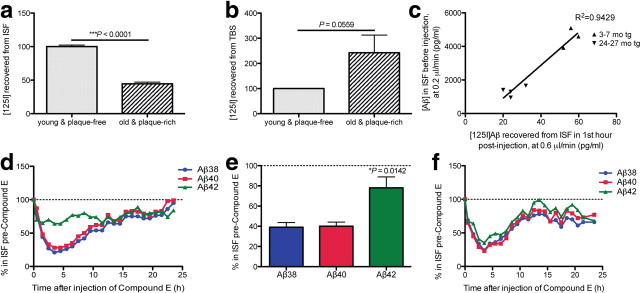
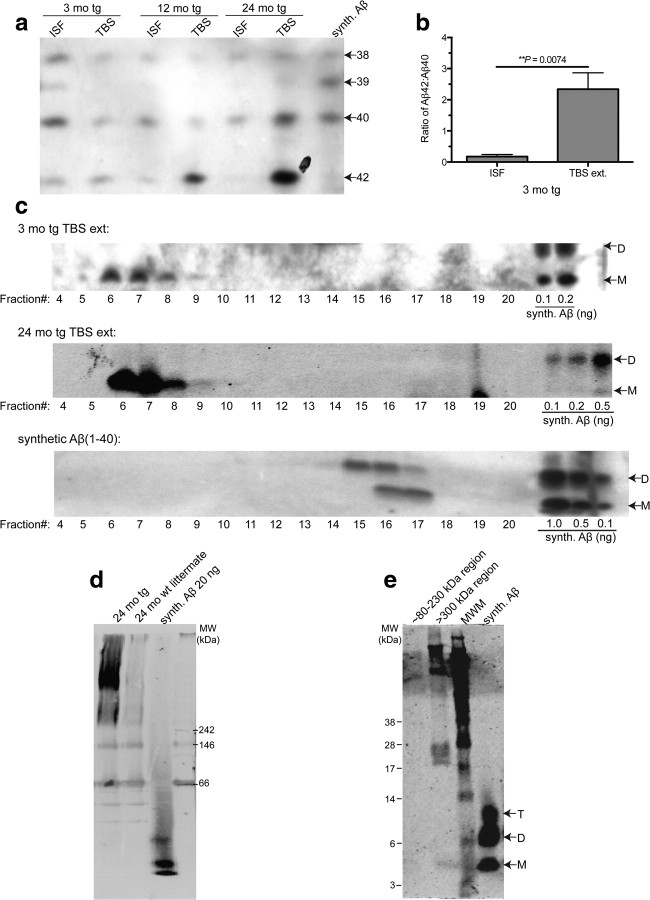
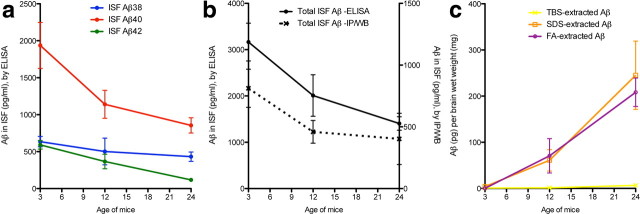
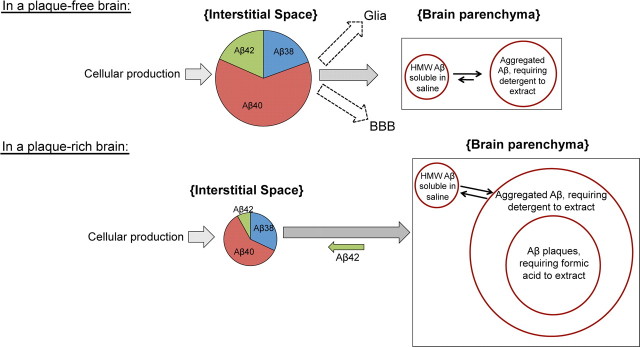
Growing evidence supports the hypothesis that soluble, diffusible forms of the amyloid β-peptide (Aβ) are pathogenically important in Alzheimer's disease (AD) and thus have both diagnostic and therapeutic salience. To learn more about the dynamics of soluble Aβ economy in vivo, we used microdialysis to sample the brain interstitial fluid (ISF), which contains the most soluble Aβ species in brain at steady state, in >40 wake, behaving APP transgenic mice before and during the process of Aβ plaque formation (age 3-28 months). Diffusible forms of Aβ, especially Aβ(42), declined significantly in ISF as mice underwent progressive parenchymal deposition of Aβ. Moreover, radiolabeled Aβ administered at physiological concentrations into ISF revealed a striking difference in the fate of soluble Aβ in plaque-rich (vs plaque-free) mice: it clears more rapidly from the ISF and becomes more associated with the TBS-extractable pool, suggesting that cerebral amyloid deposits can rapidly sequester soluble Aβ from the ISF. Likewise, acute γ-secretase inhibition in plaque-free mice showed a marked decline of Aβ(38), Aβ(40), and Aβ(42), whereas in plaque-rich mice, Aβ(42) declined significantly less. These results suggest that most of the Aβ(42) that populates the ISF in plaque-rich mice is derived not from new Aβ biosynthesis but rather from the large reservoir of less soluble Aβ(42) in brain parenchyma. Together, these and other findings herein illuminate the in vivo dynamics of soluble Aβ during the development of AD-type neuropathology and after γ-secretase inhibition and help explain the apparent paradox that CSF Aβ(42) levels fall as humans develop AD.
Figures
References
-
- Bech RA, Waldemar G, Gjerris F, Klinken L, Juhler M. Shunting effects in patients with idiopathic normal pressure hydrocephalus; correlation with cerebral and leptomeningeal biopsy findings. Acta Neurochir (Wien) 1999;141:633–639. - PubMed
-
- Cirrito JR, May PC, O'Dell MA, Taylor JW, Parsadanian M, Cramer JW, Audia JE, Nissen JS, Bales KR, Paul SM, DeMattos RB, Holtzman DM. In vivo assessment of brain interstitial fluid with microdialysis reveals plaque-associated changes in amyloid-beta metabolism and half-life. J Neurosci. 2003;23:8844–8853. - PMC - PubMed
-
- Del Bigio MR, Cardoso ER, Halliday WC. Neuropathological changes in chronic adult hydrocephalus: cortical biopsies and autopsy findings. Can J Neurol Sci. 1997;24:121–126. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases