

Upregulation of Cyclin B1 by miRNA and its implications in cancer
- PMID: 22053081
- PMCID: PMC3287204
- DOI: 10.1093/nar/gkr934
Upregulation of Cyclin B1 by miRNA and its implications in cancer
Abstract
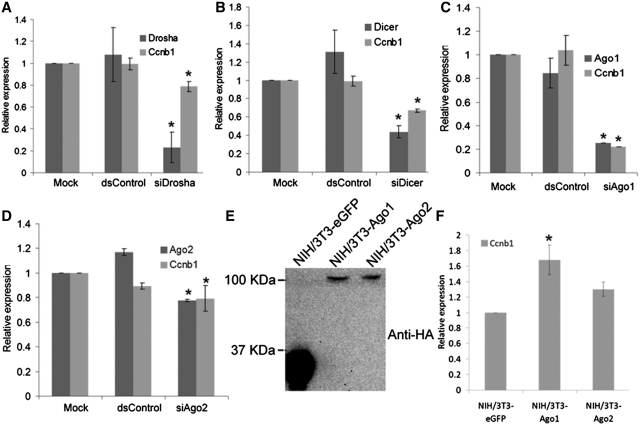
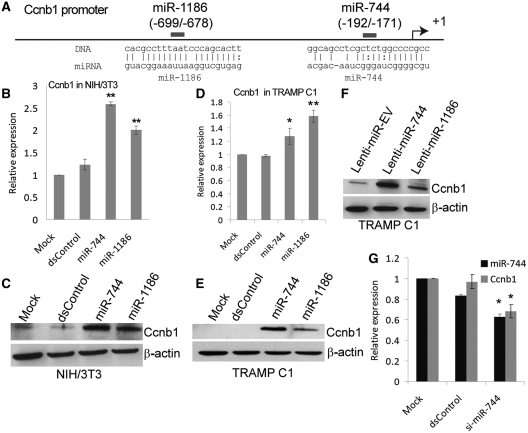
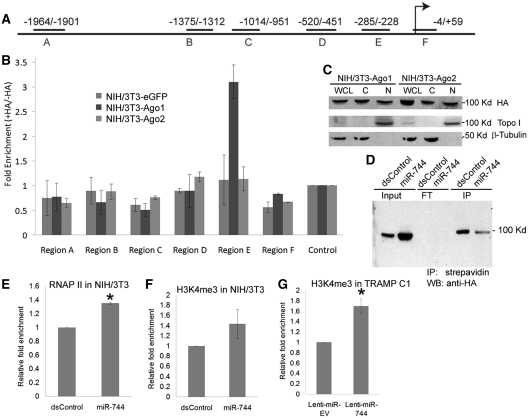
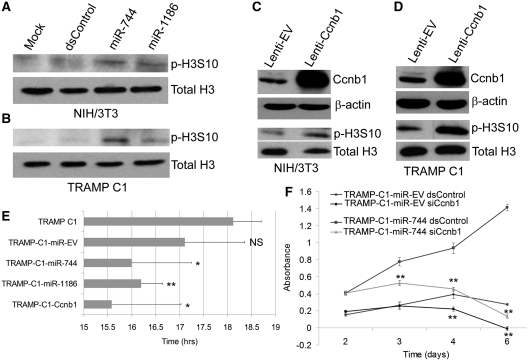
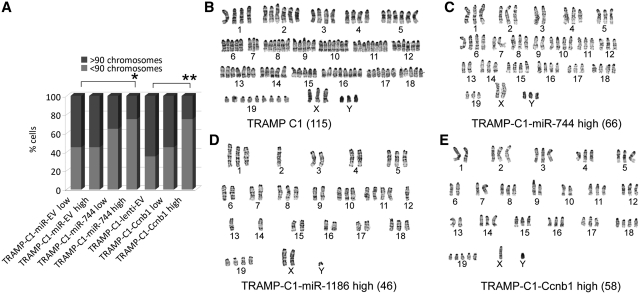
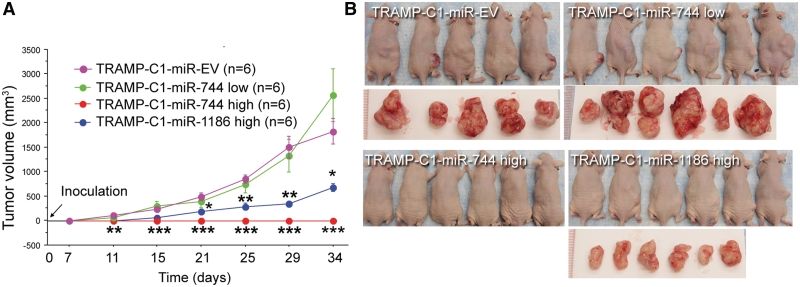
It is largely recognized that microRNAs (miRNAs) function to silence gene expression by targeting 3'UTR regions. However, miRNAs have also been implicated to positively-regulate gene expression by targeting promoter elements, a phenomenon known as RNA activation (RNAa). In the present study, we show that expression of mouse Cyclin B1 (Ccnb1) is dependent on key factors involved in miRNA biogenesis and function (i.e. Dicer, Drosha, Ago1 and Ago2). In silico analysis identifies highly-complementary sites for 21 miRNAs in the Ccnb1 promoter. Experimental validation identified three miRNAs (miR-744, miR-1186 and miR-466d-3p) that induce Ccnb1 expression in mouse cell lines. Conversely, knockdown of endogenous miR-744 led to decreased Ccnb1 levels. Chromatin immunoprecipitation (ChIP) analysis revealed that Ago1 was selectively associated with the Ccnb1 promoter and miR-744 increased enrichment of RNA polymerase II (RNAP II) and trimethylation of histone 3 at lysine 4 (H3K4me3) at the Ccnb1 transcription start site. Functionally, short-term overexpression of miR-744 and miR-1186 resulted in enhanced cell proliferation, while prolonged expression caused chromosomal instability and in vivo tumor suppression. Such phenotypes were recapitulated by overexpression of Ccnb1. Our findings reveal an endogenous system by which miRNA functions to activate Ccnb1 expression in mouse cells and manipulate in vivo tumor development/growth.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
