

Comprehensive survey and geometric classification of base triples in RNA structures
- PMID: 22053086
- PMCID: PMC3287178
- DOI: 10.1093/nar/gkr810
Comprehensive survey and geometric classification of base triples in RNA structures
Abstract
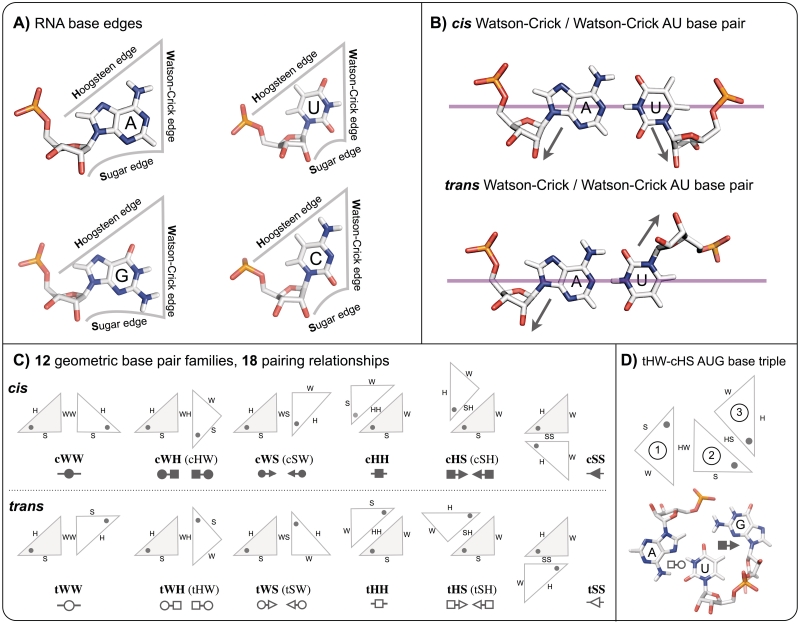
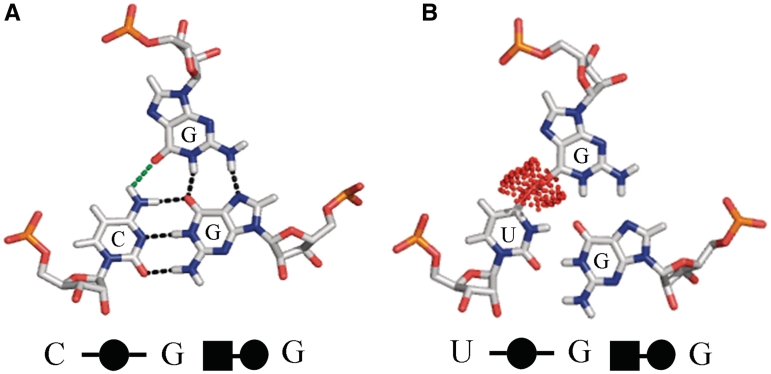
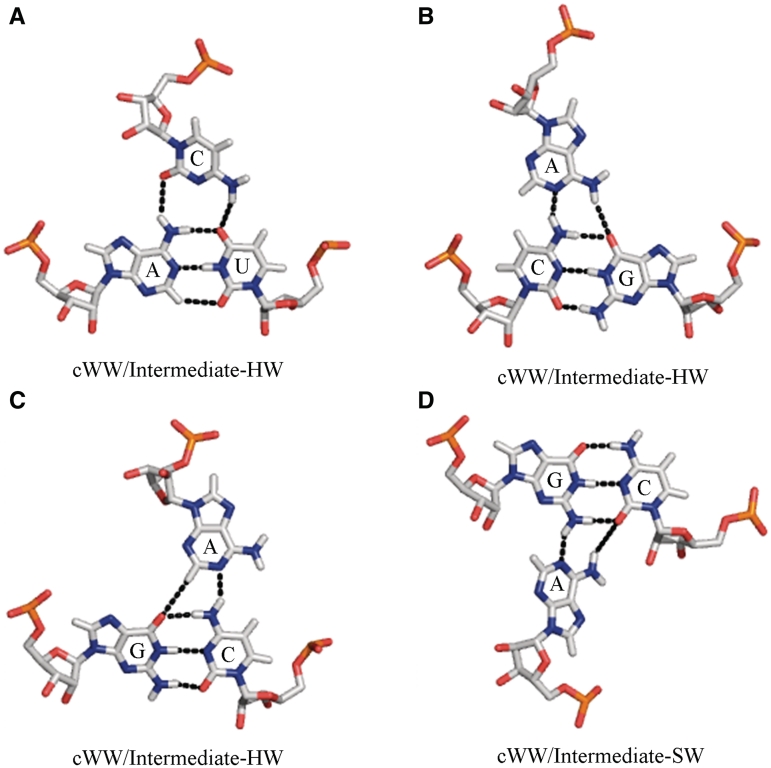
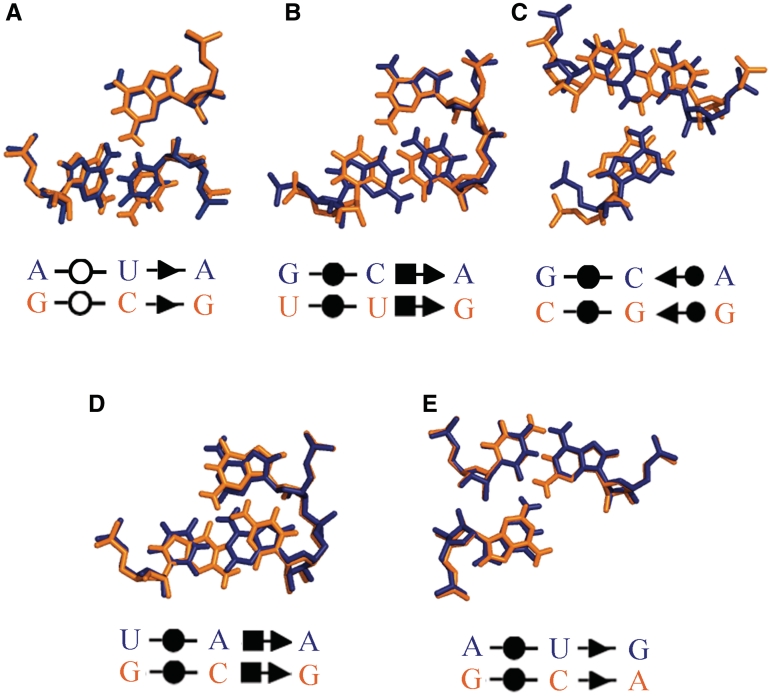
Base triples are recurrent clusters of three RNA nucleobases interacting edge-to-edge by hydrogen bonding. We find that the central base in almost all triples forms base pairs with the other two bases of the triple, providing a natural way to geometrically classify base triples. Given 12 geometric base pair families defined by the Leontis-Westhof nomenclature, combinatoric enumeration predicts 108 potential geometric base triple families. We searched representative atomic-resolution RNA 3D structures and found instances of 68 of the 108 predicted base triple families. Model building suggests that some of the remaining 40 families may be unlikely to form for steric reasons. We developed an on-line resource that provides exemplars of all base triples observed in the structure database and models for unobserved, predicted triples, grouped by triple family, as well as by three-base combination (http://rna.bgsu.edu/Triples). The classification helps to identify recurrent triple motifs that can substitute for each other while conserving RNA 3D structure, with applications in RNA 3D structure prediction and analysis of RNA sequence evolution.
Figures
References
-
- Leontis NB, Westhof E. A common motif organizes the structure of multi-helix loops in 16 S and 23 S ribosomal RNAs. J. Mol. Biol. 1998;283:571–583. - PubMed
-
- Szewczak AA, Moore PB. The sarcin/ricin loop, a modular RNA. J. Mol. Biol. 1995;247:81–98. - PubMed
-
- Doherty EA, Batey RT, Masquida B, Doudna JA. A universal mode of helix packing in RNA. Nat. Struct. Biol. 2001;8:339–343. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
