

The 3'-to-5' exoribonuclease Nibbler shapes the 3' ends of microRNAs bound to Drosophila Argonaute1
- PMID: 22055293
- PMCID: PMC3236499
- DOI: 10.1016/j.cub.2011.09.034
The 3'-to-5' exoribonuclease Nibbler shapes the 3' ends of microRNAs bound to Drosophila Argonaute1
Abstract
Background: MicroRNAs (miRNAs) are ~22 nucleotide (nt) small RNAs that control development, physiology, and pathology in animals and plants. Production of miRNAs involves the sequential processing of primary hairpin-containing RNA polymerase II transcripts by the RNase III enzymes Drosha in the nucleus and Dicer in the cytoplasm. miRNA duplexes then assemble into Argonaute proteins to form the RNA-induced silencing complex (RISC). In mature RISC, a single-stranded miRNA directs the Argonaute protein to bind partially complementary sequences, typically in the 3' untranslated regions of messenger RNAs, repressing their expression.
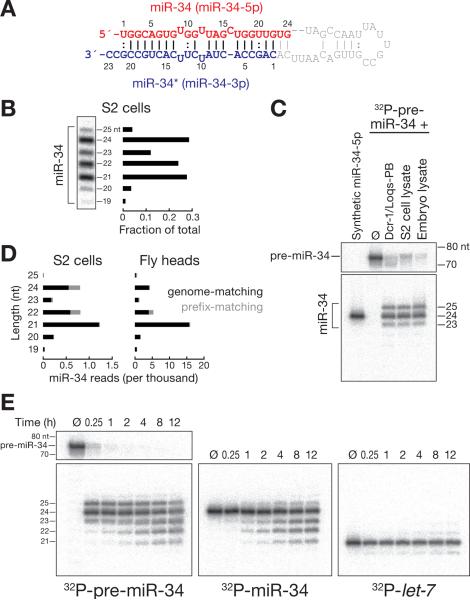
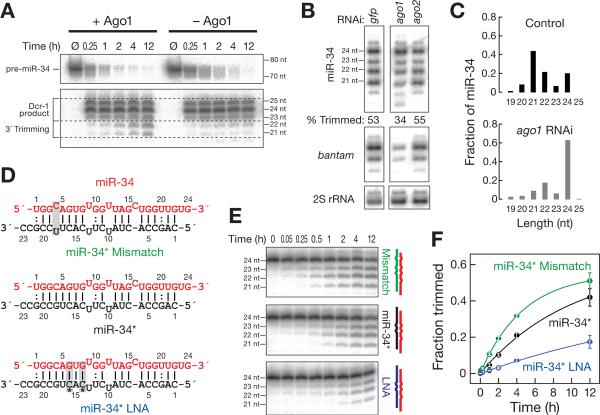
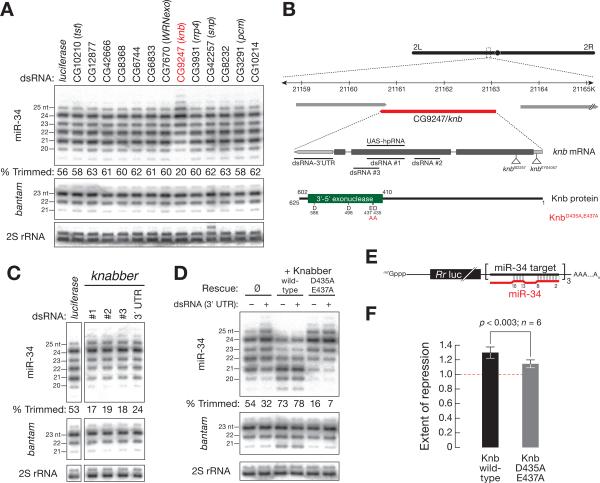
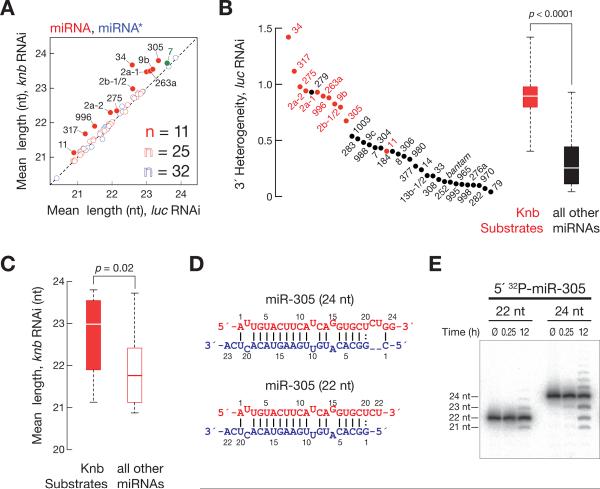
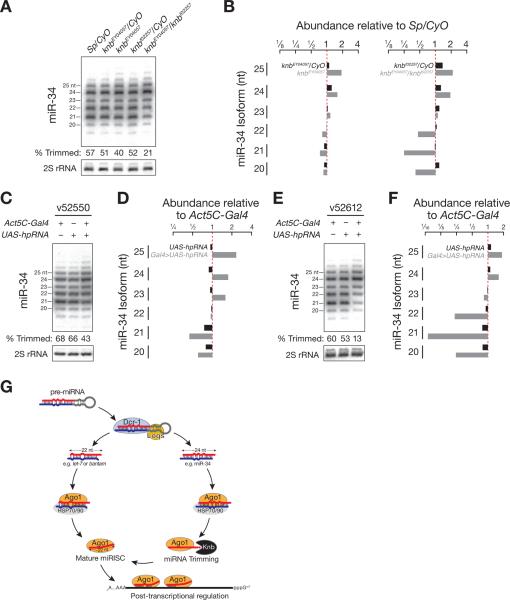
Results: Here, we show that after loading into Argonaute1 (Ago1), more than a quarter of all Drosophila miRNAs undergo 3' end trimming by the 3'-to-5' exoribonuclease Nibbler (CG9247). Depletion of Nibbler by RNA interference (RNAi) reveals that miRNAs are frequently produced by Dicer-1 as intermediates that are longer than ~22 nt. Trimming of miRNA 3' ends occurs after removal of the miRNA* strand from pre-RISC and may be the final step in RISC assembly, ultimately enhancing target messenger RNA repression. In vivo, depletion of Nibbler by RNAi causes developmental defects.
Conclusions: We provide a molecular explanation for the previously reported heterogeneity of miRNA 3' ends and propose a model in which Nibbler converts miRNAs into isoforms that are compatible with the preferred length of Ago1-bound small RNAs.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
References
-
- Filipowicz W, Bhattacharyya SN, Sonenberg N. Mechanisms of post-transcriptional regulation by microRNAs: are the answers in sight? Nat Rev Genet. 2008;9:102–114. - PubMed
-
- Lee RC, Feinbaum RL, Ambros V. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-14. Cell. 1993;75:843–854. - PubMed
-
- Wightman B, Ha I, Ruvkun G. Posttranscriptional regulation of the heterochronic gene lin-14 by lin-4 mediates temporal pattern formation in C. elegans. Cell. 1993;75:855–862. - PubMed
-
- Bushati N, Cohen SM. microRNA functions. Annu Rev Cell Dev Biol. 2007;23:175–205. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
