

Dissociation of the glucose and lipid regulatory functions of FoxO1 by targeted knockin of acetylation-defective alleles in mice
- PMID: 22055502
- PMCID: PMC3221516
- DOI: 10.1016/j.cmet.2011.09.012
Dissociation of the glucose and lipid regulatory functions of FoxO1 by targeted knockin of acetylation-defective alleles in mice
Abstract
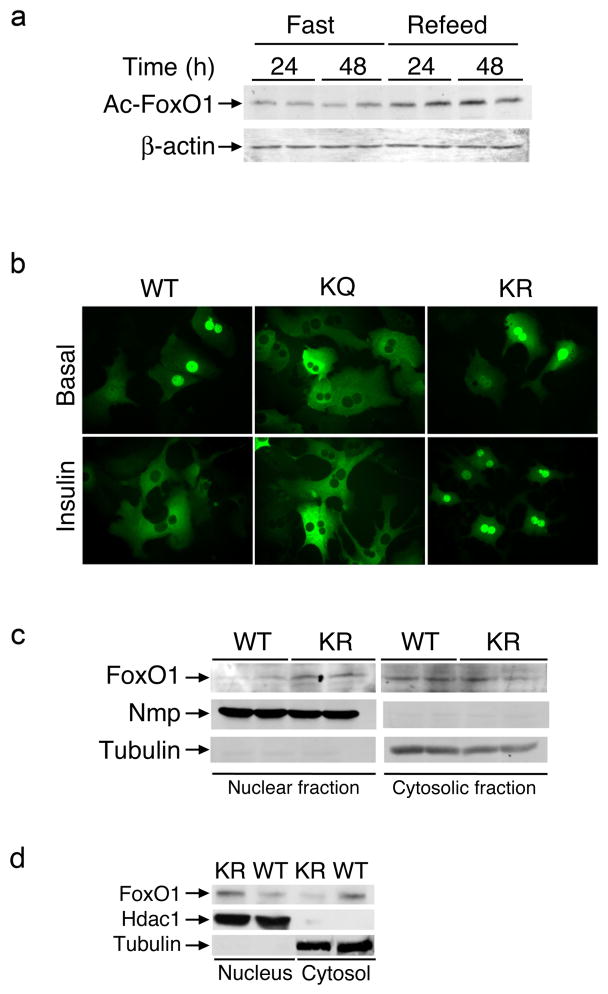
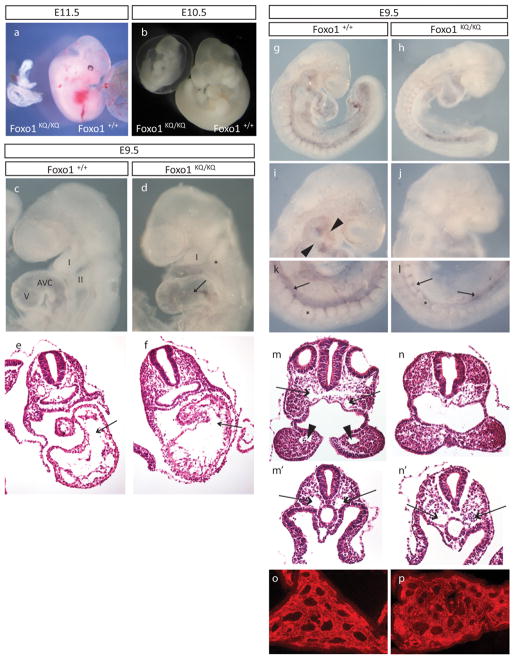
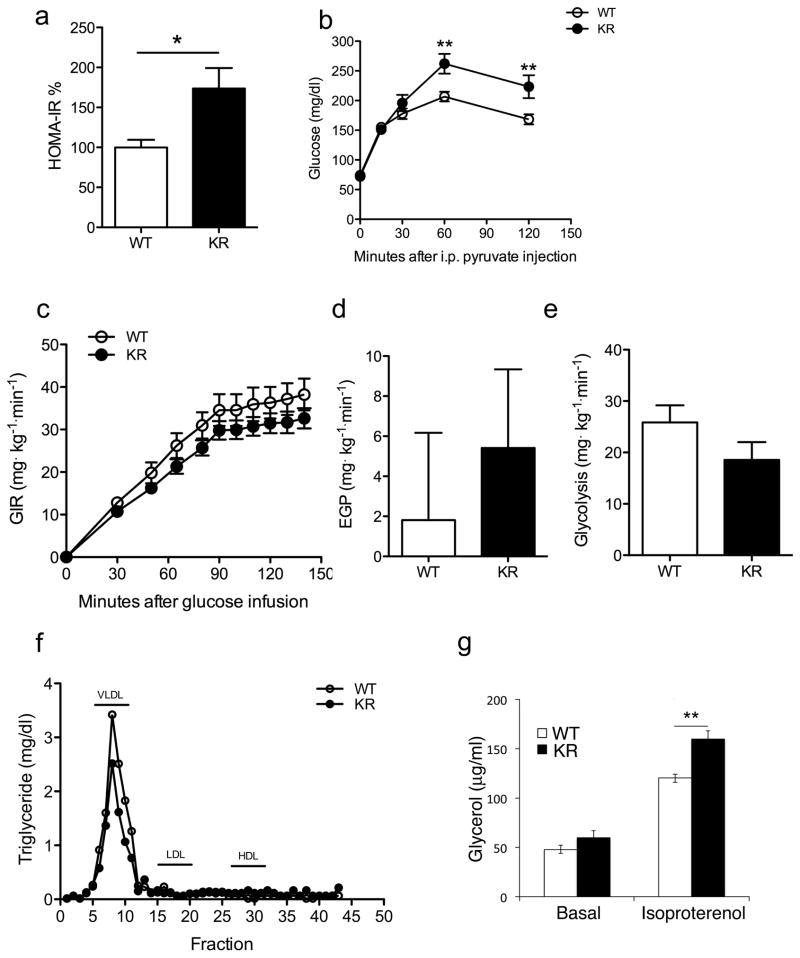
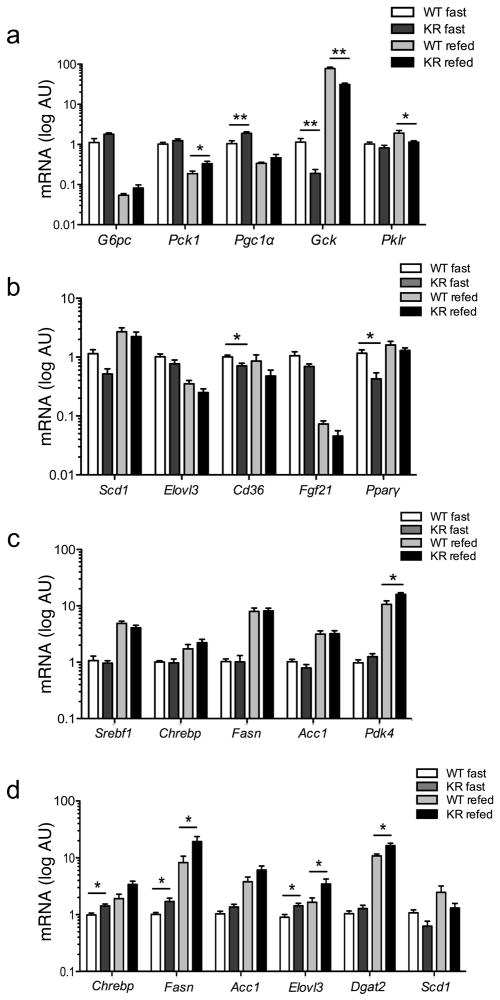
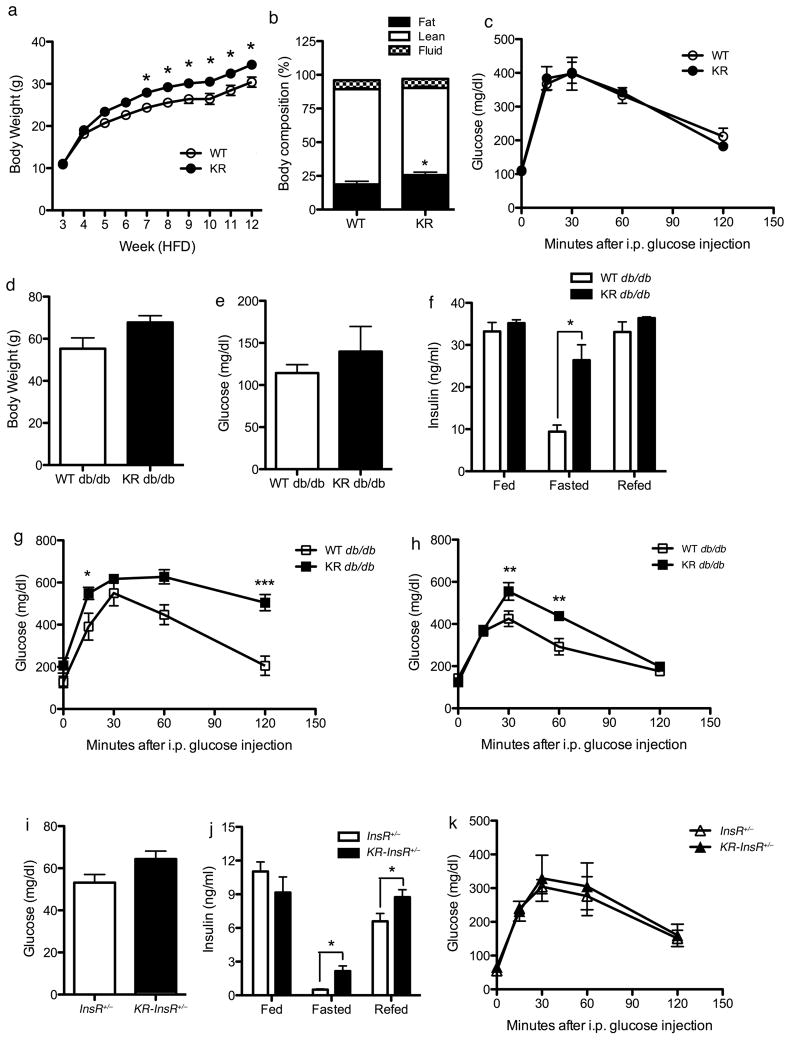
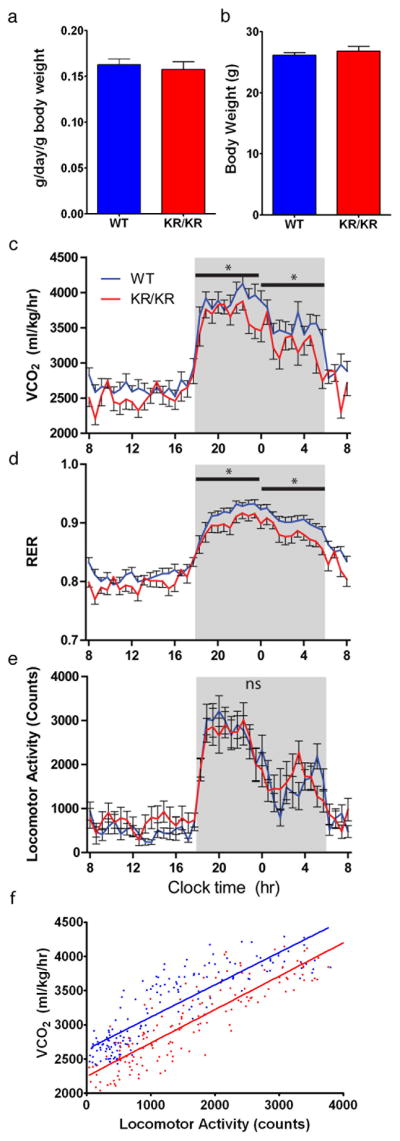
FoxO1 integrates multiple metabolic pathways. Nutrient levels modulate FoxO1 acetylation, but the functional consequences of this posttranslational modification are unclear. To answer this question, we generated mice bearing alleles that encode constitutively acetylated and acetylation-defective FoxO1 proteins. Homozygosity for an allele mimicking constitutive acetylation (Foxo1(KQ/KQ)) results in embryonic lethality due to cardiac and angiogenesis defects. In contrast, mice homozygous for a constitutively deacetylated Foxo1 allele (Foxo1(KR/KR)) display a unique metabolic phenotype of impaired insulin action on hepatic glucose metabolism but decreased plasma lipid levels and low respiratory quotient that are consistent with a state of preferential lipid usage. Moreover, Foxo1(KR/KR) mice show a dissociation between weight gain and insulin resistance in predisposing conditions (high fat diet, diabetes, and insulin receptor mutations), possibly due to decreased cytokine production in adipose tissue. Thus, acetylation inactivates FoxO1 during nutrient excess whereas deacetylation selectively potentiates FoxO1 activity, protecting against excessive catabolism during nutrient deprivation.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
References
-
- Accili D, Arden KC. FoxOs at the crossroads of cellular metabolism, differentiation, and transformation. Cell. 2004;117:421–426. - PubMed
-
- Accili D, Drago J, Lee EJ, Johnson MD, Cool MH, Salvatore P, Asico LD, Jose PA, Taylor SI, Westphal H. Early neonatal death in mice homozygous for a null allele of the insulin receptor gene. Nat Genet. 1996;12:106–109. - PubMed
-
- Allen-Jennings AE, Hartman MG, Kociba GJ, Hai T. The roles of atf3 in glucose homeostasis. a transgenic mouse model with liver dysfunction and defects in endocrine pancreas. J Biol Chem. 2001;276:29507–29514. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- F32 DK079496/DK/NIDDK NIH HHS/United States
- U24 DK059635/DK/NIDDK NIH HHS/United States
- HL087123/HL/NHLBI NIH HHS/United States
- R01 DK040936/DK/NIDDK NIH HHS/United States
- R37 DK058282/DK/NIDDK NIH HHS/United States
- R01 DK064819/DK/NIDDK NIH HHS/United States
- R01 DK057539/DK/NIDDK NIH HHS/United States
- P30 DK063608/DK/NIDDK NIH HHS/United States
- DK63608/DK/NIDDK NIH HHS/United States
- DK079496/DK/NIDDK NIH HHS/United States
- DK057539/DK/NIDDK NIH HHS/United States
- U24 DK076169/DK/NIDDK NIH HHS/United States
- P01 HL087123/HL/NHLBI NIH HHS/United States
- DK059635/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
