

Formins filter modified actin subunits during processive elongation
- PMID: 22056467
- PMCID: PMC3683246
- DOI: 10.1016/j.jsb.2011.10.005
Formins filter modified actin subunits during processive elongation
Abstract
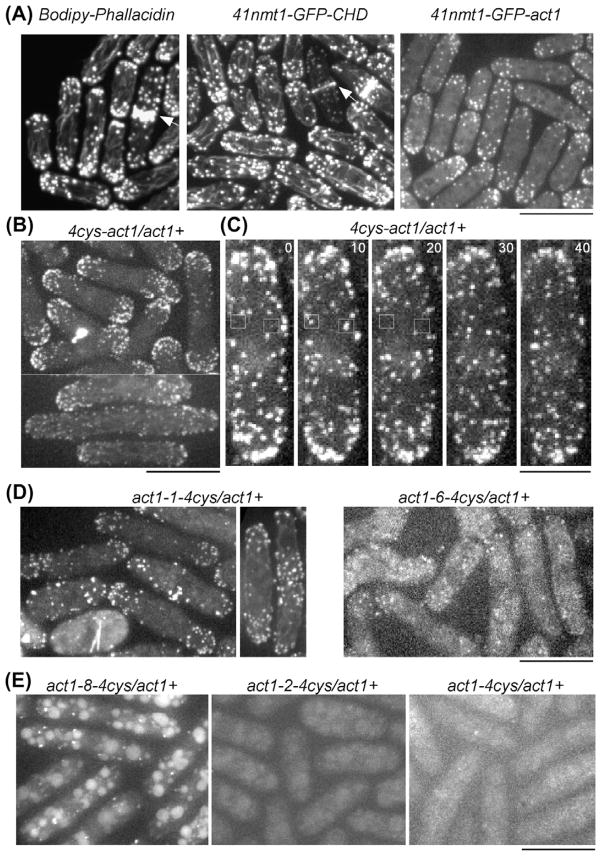
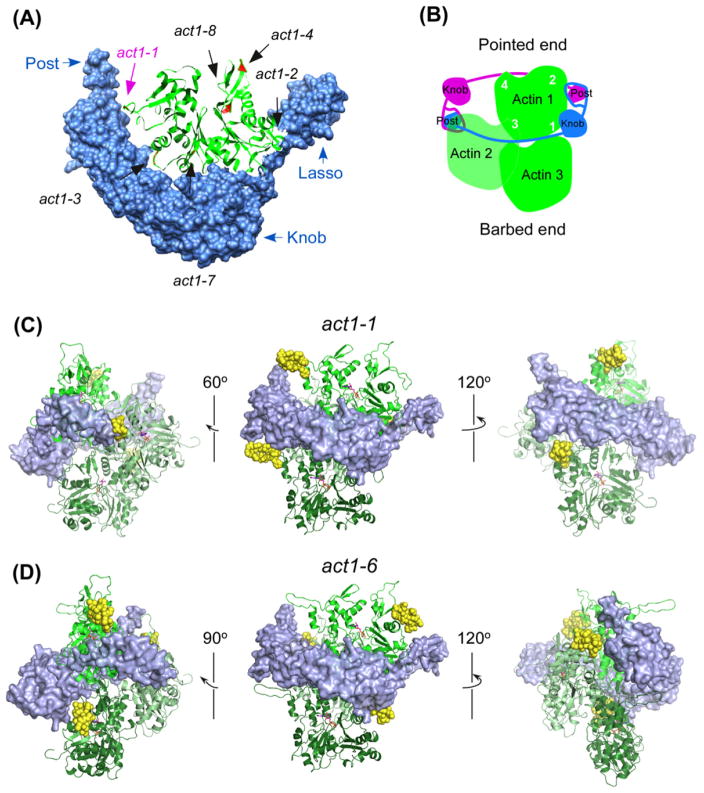
Fission yeast cells reject actin subunits tagged with a fluorescent protein from the cytokinetic contractile ring, so cytokinesis fails and the cells die when the native actin gene is replaced by GFP-actin. The lack of a fluorescent actin probe has prevented a detailed study of actin filament dynamics in contractile rings, and left open questions regarding the mechanism of cytokinesis. To incorporate fluorescent actin into the contractile ring to study its dynamics, we introduced the coding sequence for a tetracysteine motif (FLNCCPGCCMEP) at 10 locations in the fission yeast actin gene and expressed the mutant proteins from the native actin locus in diploid cells with wild-type actin on the other chromosome. We labeled these tagged actins inside live cells with the FlAsH reagent. Cells incorporated some of these labeled actins into actin patches at sites of endocytosis, where Arp2/3 complex nucleates all of the actin filaments. However, the cells did not incorporate any of the FlAsH-actins into the contractile ring. Therefore, formin Cdc12p rejects actin subunits with a tag of ~2 kDa, illustrating the stringent structural requirements for this formin to promote the elongation of actin filament barbed ends as it moves processively along the end of a growing filament.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures

References
-
- Adams SR, Campbell RE, Gross LA, Martin BR, Walkup GK, et al. New biarsenical ligands and tetracysteine motifs for protein labeling in vitro and in vivo: synthesis and biological applications. J Am Chem Soc. 2002;124:6063–6076. - PubMed
-
- Aebi U, Smith PR, Isenberg G, Pollard TD. Structure of crystalline actin sheets. Nature. 1980;288:296–298. - PubMed
-
- Aizawa H, Sameshima M, Yahara I. A green fluorescent protein-actin fusion protein dominantly inhibits cytokinesis, cell spreading, and locomotion in Dictyostelium. Cell Struct Funct. 1997;22:335–345. - PubMed
-
- Arnold K, Bordoli L, Kopp J, Schwede T. The SWISS-MODEL workspace: a web-based environment for protein structure homology modelling. Bioinformatics. 2006;22:195–201. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
