

Rejuvenating senescent and centenarian human cells by reprogramming through the pluripotent state
- PMID: 22056670
- PMCID: PMC3219229
- DOI: 10.1101/gad.173922.111
Rejuvenating senescent and centenarian human cells by reprogramming through the pluripotent state
Abstract
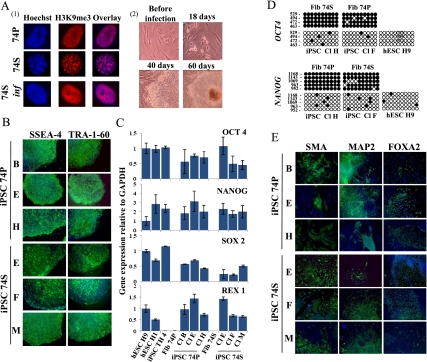
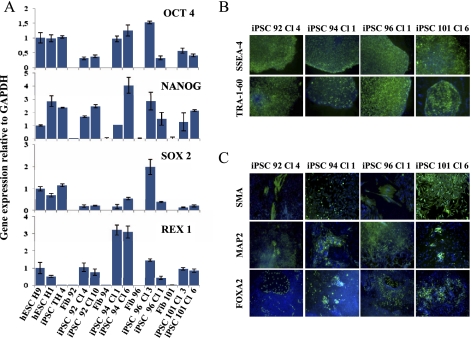
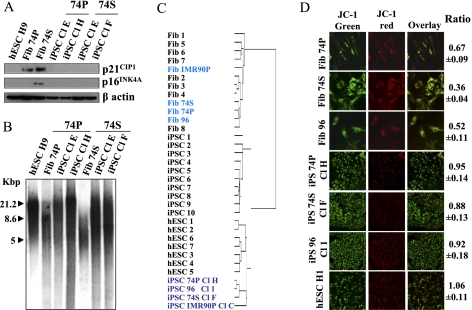
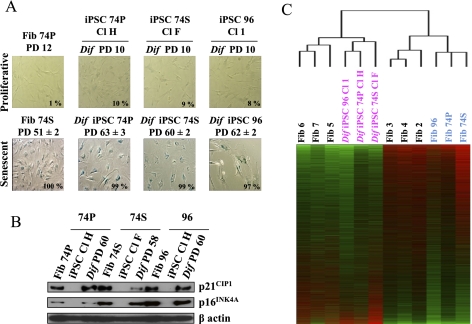
Direct reprogramming of somatic cells into induced pluripotent stem cells (iPSCs) provides a unique opportunity to derive patient-specific stem cells with potential applications in tissue replacement therapies and without the ethical concerns of human embryonic stem cells (hESCs). However, cellular senescence, which contributes to aging and restricted longevity, has been described as a barrier to the derivation of iPSCs. Here we demonstrate, using an optimized protocol, that cellular senescence is not a limit to reprogramming and that age-related cellular physiology is reversible. Thus, we show that our iPSCs generated from senescent and centenarian cells have reset telomere size, gene expression profiles, oxidative stress, and mitochondrial metabolism, and are indistinguishable from hESCs. Finally, we show that senescent and centenarian-derived pluripotent stem cells are able to redifferentiate into fully rejuvenated cells. These results provide new insights into iPSC technology and pave the way for regenerative medicine for aged patients.
Figures
References
-
- Alexander B, Coppola G, Perrault SD, Peura TT, Betts DH, King WA 2007. Telomere length status of somatic cell sheep clones and their offspring. Mol Reprod Dev 74: 1525–1537 - PubMed
-
- Armstrong L, Tilgner K, Saretzki G, Atkinson SP, Stojkovic M, Moreno R, Przyborski S, Lako M 2010. Human induced pluripotent stem cell lines show stress defense mechanisms and mitochondrial regulation similar to those of human embryonic stem cells. Stem Cells 28: 661–673 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases