

A dynamical systems analysis of afferent control in a neuromechanical model of locomotion: II. Phase asymmetry
- PMID: 22058275
- PMCID: PMC3422648
- DOI: 10.1088/1741-2560/8/6/065004
A dynamical systems analysis of afferent control in a neuromechanical model of locomotion: II. Phase asymmetry
Abstract
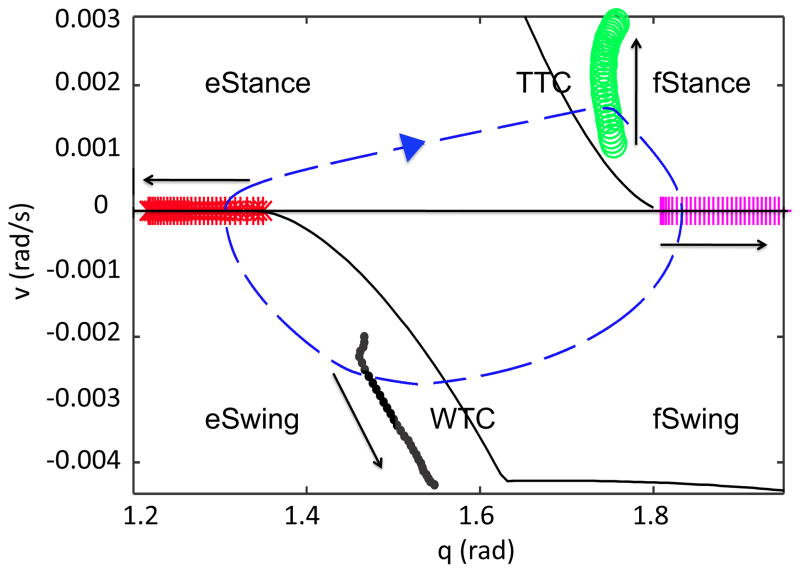
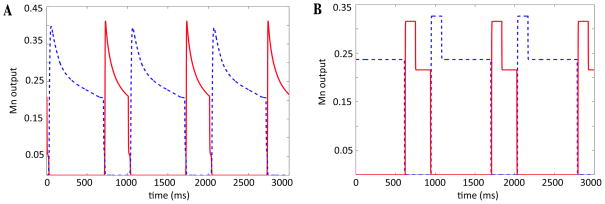
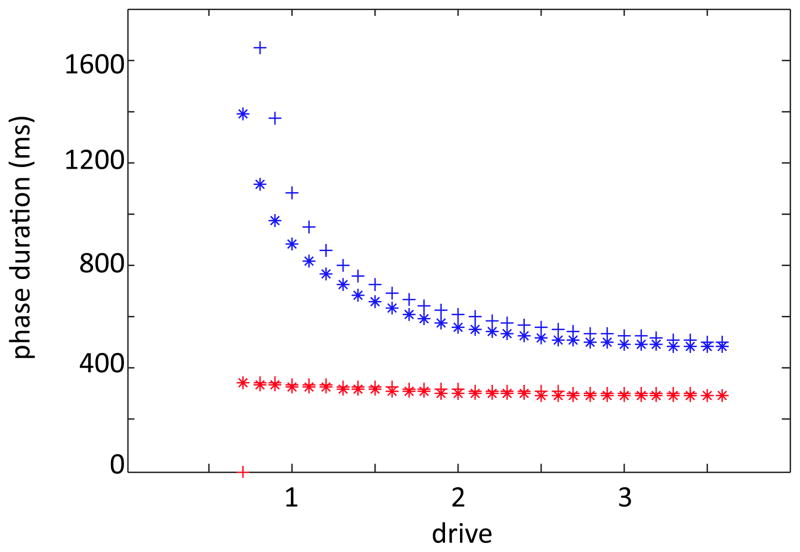
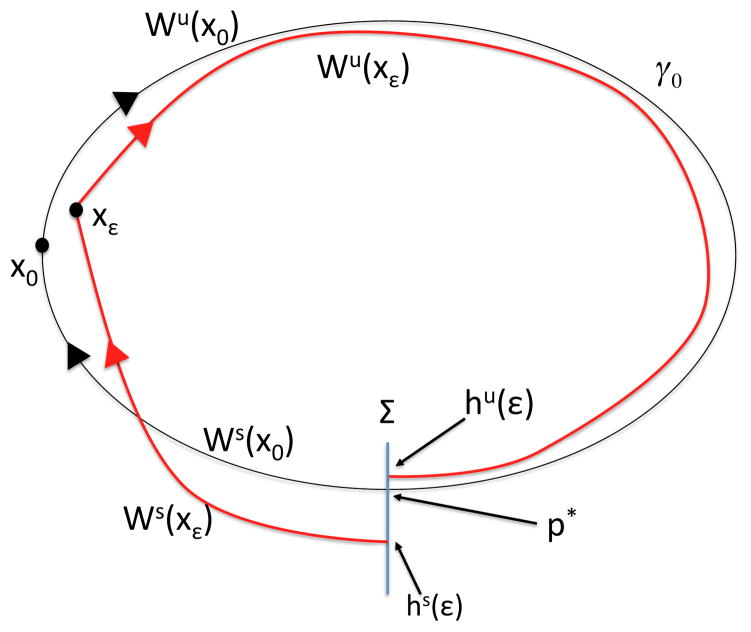
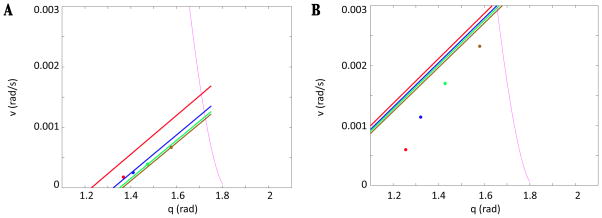
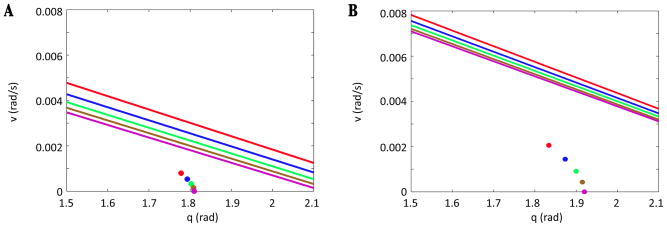
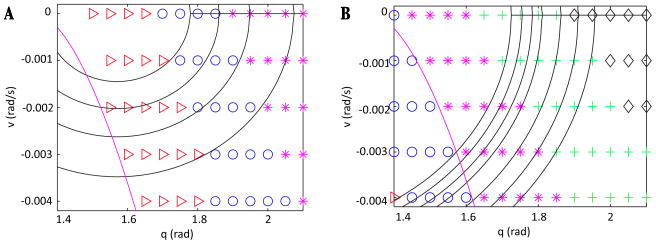
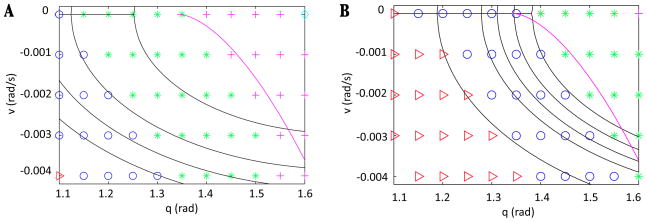
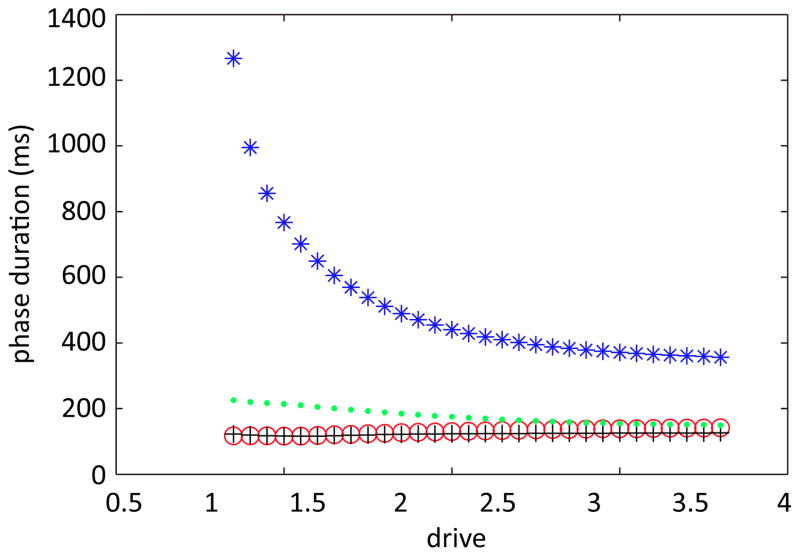
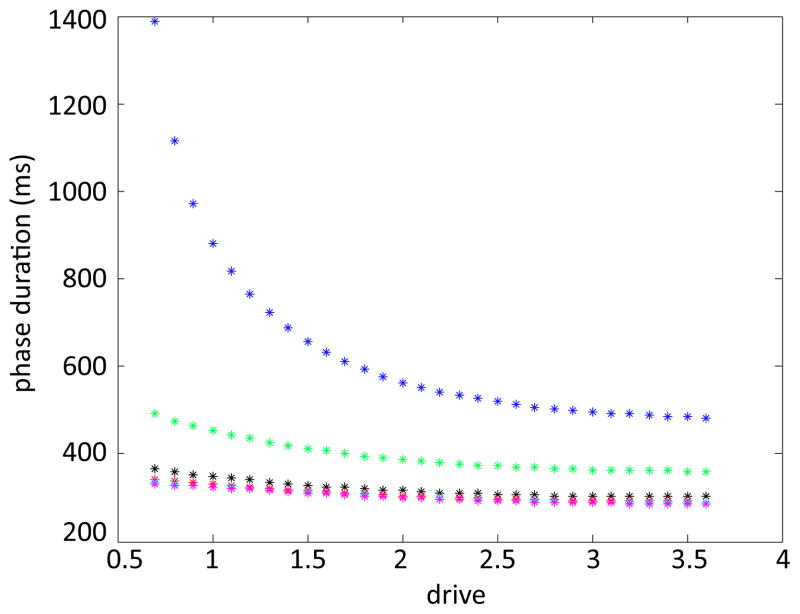
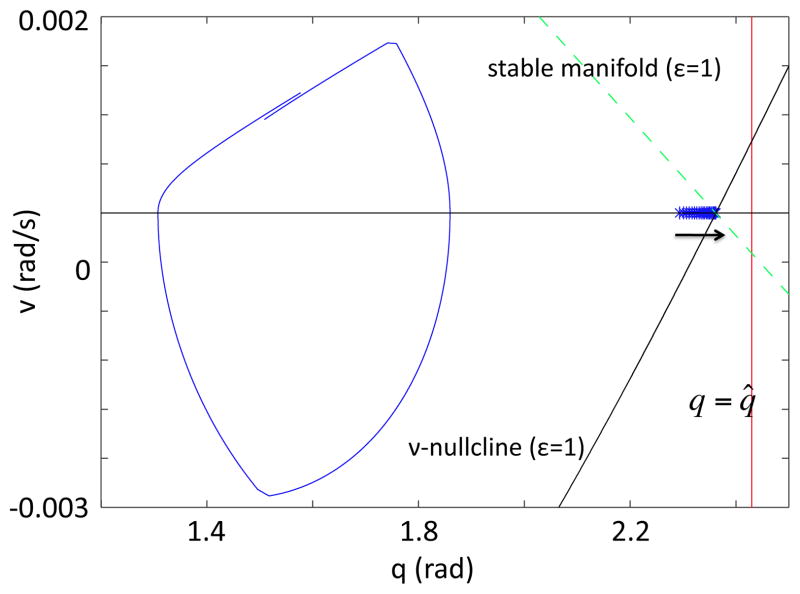
In this paper we analyze a closed loop neuromechanical model of locomotor rhythm generation. The model is composed of a spinal central pattern generator (CPG) and a single-joint limb, with CPG outputs projecting via motoneurons to muscles that control the limb and afferent signals from the muscles feeding back to the CPG. In a preceding companion paper (Spardy et al 2011 J. Neural Eng. 8 065003), we analyzed how the model generates oscillations in the presence or absence of feedback, identified curves in a phase plane associated with the limb that signify where feedback levels induce phase transitions within the CPG, and explained how increasing feedback strength restores oscillations in a model representation of spinal cord injury; from these steps, we derived insights about features of locomotor rhythms in several scenarios and made predictions about rhythm responses to various perturbations. In this paper, we exploit our analytical observations to construct a reduced model that retains important characteristics from the original system. We prove the existence of an oscillatory solution to the reduced model using a novel version of a Melnikov function, adapted for discontinuous systems, and also comment on the uniqueness and stability of this solution. Our analysis yields a deeper understanding of how the model must be tuned to generate oscillations and how the details of the limb dynamics shape overall model behavior. In particular, we explain how, due to the feedback signals in the model, changes in the strength of a tonic supra-spinal drive to the CPG yield asymmetric alterations in the durations of different locomotor phases, despite symmetry within the CPG itself.
Figures
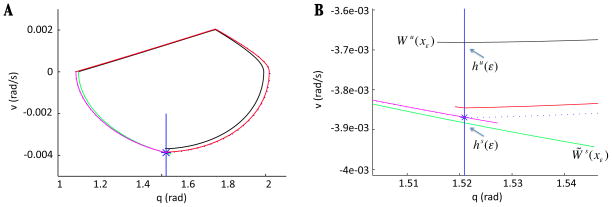
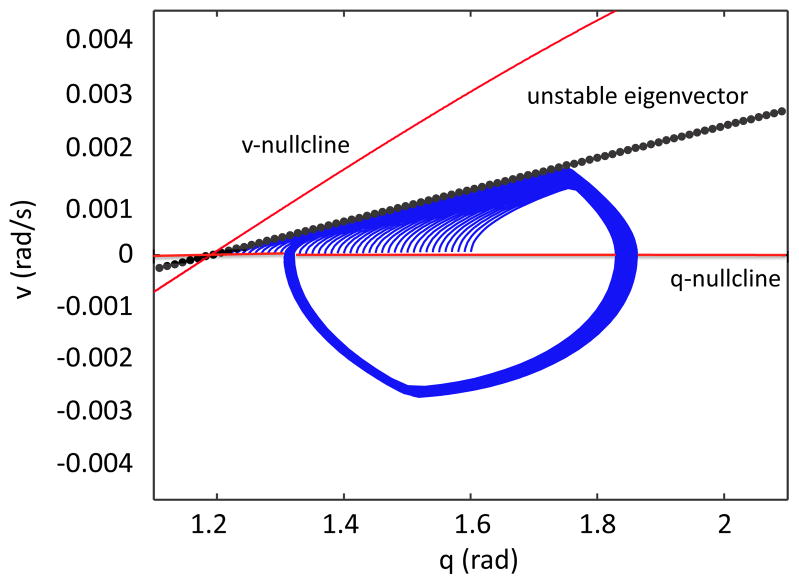
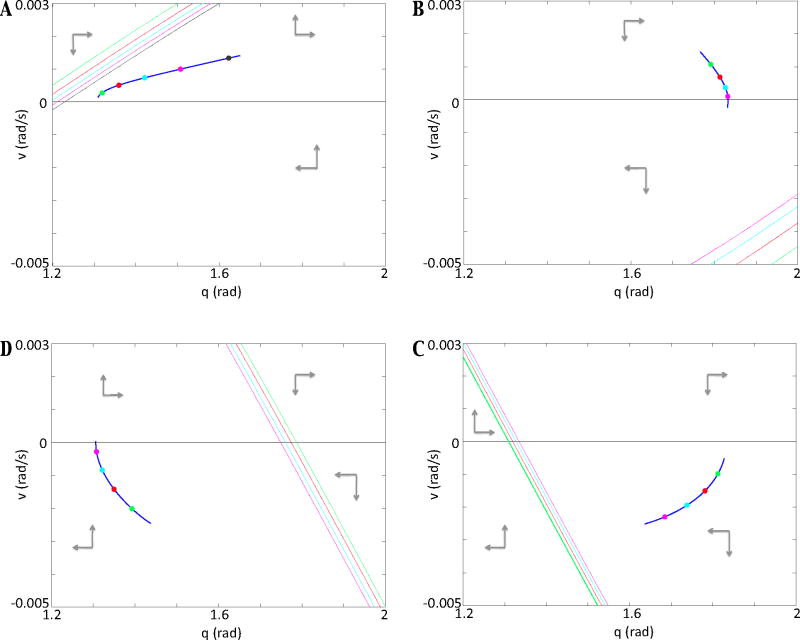
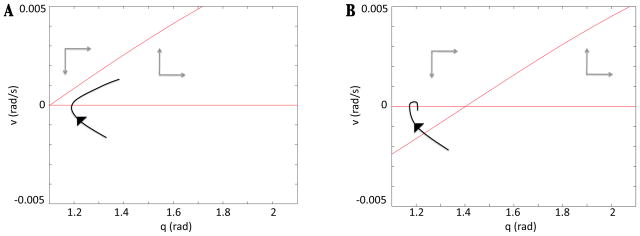
References
-
- Brown TG. The intrinsic factors in the act of progression in the mammal. Proceedings of the Royal Society of London. Series B, Containing Papers of a Biological Character. 1911;84(572):308–319.
-
- Jankowska E, Jukes MGM, Lund S, Lundberg A. The effect of dopa on the spinal cord 6. half-centre organization of interneurones transmitting effects from the flexor reflex afferents. Acta Physiol Scand. 1967;70(3–4):389–402. - PubMed
-
- Lundberg JANM, ”Anggård A, Fahrenkrug Jan. Complementary role of vasoactive intestinal polypeptide (vip) and acetylcholine for cat submandibular gland blood flow and secretion i. vip release. Acta Physiol Scand. 1981;113(3):317–327. - PubMed
-
- Lafreniere-Roula M, McCrea DA. Deletions of rhythmic motoneuron activity during fictive locomotion and scratch provide clues to the organization of the mammalian central pattern generator. J Neurophysiol. 2005;94(2):1120. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources