

Review
doi: 10.1016/j.bbamcr.2011.10.009.
Epub 2011 Oct 28.
The mechanism of dynein motility: insight from crystal structures of the motor domain
Affiliations
- PMID: 22062687
- PMCID: PMC3249483
- DOI: 10.1016/j.bbamcr.2011.10.009
Item in Clipboard
Review
The mechanism of dynein motility: insight from crystal structures of the motor domain
Biochim Biophys Acta.
2012 Jan.
Abstract
Dynein is a large cytoskeletal motor protein that belongs to the AAA+ (ATPases associated with diverse cellular activities) superfamily. While dynein has had a rich history of cellular research, its molecular mechanism of motility remains poorly understood. Here we describe recent X-ray crystallographic studies that reveal the architecture of dynein's catalytic ring, mechanical linker element, and microtubule binding domain. This structural information has given rise to new hypotheses on how the dynein motor domain might change its conformation in order to produce motility along microtubules.
Copyright © 2011 Elsevier B.V. All rights reserved.
Figures
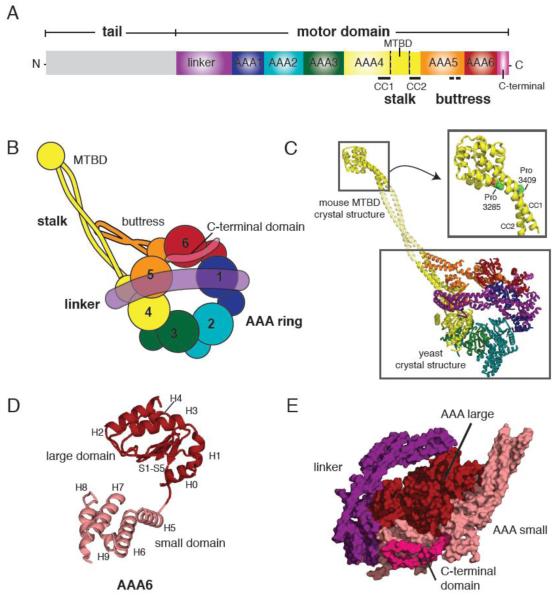
(A) Primary structure of the dynein heavy chain showing the functional domains of dynein. (MTBD = microtubule binding domain, CC=coiled coil) (B) Cartoon depiction of the dynein motor domain as revealed by the crystal structures of yeast and Dictyostelium cytoplasmic dynein. (C) A crystallographic model of yeast cytoplasmic dynein based on the 6 Å crystal structure of the motor domain (lower box, PDB ID: 3QMZ) and the 2.3 Å resolution structure of the mouse cytoplasmic dynein MTBD (upper box, PDB ID: 3ERR). Domains are color-coded as in (B). Inset shows the MTBD structure in detail, highlighting the proline residues in CC1 and CC2 that induce a kink in the stalk. The middle portion of the stalk coiled coil is modeled from a typical anti-parallel coiled coil. (D) Architecture of the yeast dynein AAA6 domain. The large domain (H0-H4 and S1-S5) is colored in red, and the small domain (H5-H9) in pink. A flexible peptide connects the large and small domains. (E) A side view of the AAA ring, linker, and C-terminal domain. Large domains (red) form one level near the linker (purple), while the small domains (pink) form another level near the C-terminal domain (magenta).
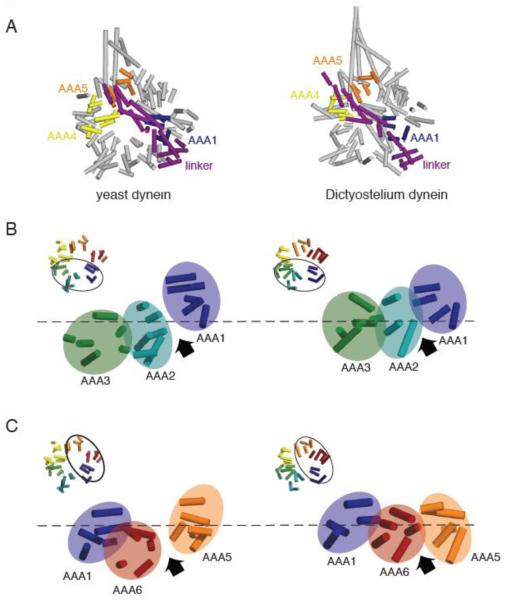
(A) Comparison of the yeast (PDB ID: 3QMZ) and Dictyostelium dynein motor domain structures (PDB ID: 3AY1). The motor domains were superimposed by aligning the helices in the linker and AAA1, which superimpose well in the yeast and Dictyostelium structures. In the yeast dynein structure, the ring is more open and asymmetric than in the Dictyostelium dynein structure. Also, AAA5 is more shifted towards AAA4 in the yeast structure, resulting in a slight difference in the position where the N-terminus of the linker sits relative to the ring. (B) Comparison of yeast and Dictyostelium dynein AAA1-3. Asymmetry is more prominent and the AAA1-2 gap is wider in the yeast structure due to the different heights of AAA domains; specifically AAA2 is shifted further down with respect to AAA1. (C) Comparison of yeast and Dicytostelium dynein AAA1, 5, and 6. The AAA ring is more planar and the AAA5-6 gap smaller in the Dictyostelium structure. As discussed in the text, we speculate that the differences between yeast and Dictyostelium reflect distinct nucleotide occupancy in AAA1 (empty and ADP for yeast and Dictyostelium respectively). Note: the model for Dictyostelium dynein lacks several AAA helices that are present in the yeast model.
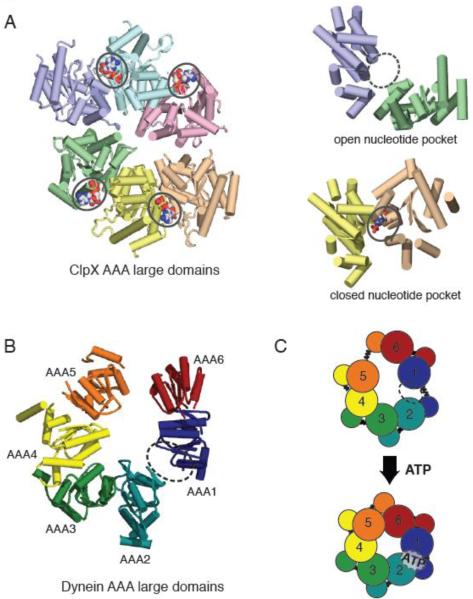
(A) X-ray crystal structure of the large domains of the ClpX hexamer (PDB ID: 3HWS), color coded by chain. The nucleotide-free domain interfaces (purple-green) show an open conformation, while the nucleotide-bound (ADP) domain interfaces (yellow-peach) show a closed conformation. (B) Asymmetric structure of the yeast cytoplasmic dynein AAA ring (only large domains shown). When viewed from the linker-face, the ring shows prominent openings between AAA1-AAA2 and AAA5-AAA6 large domains. The AAA1-AAA2 interface is the main ATP hydrolysis site for dynein. (C) A model for AAA ring communication in dynein, based upon the nucleotide-free and bound forms of ClpX. Upon ATP binding at the AAA1 nucleotide binding pocket, the AAA1-AAA2 switches from an open to closed conformation, triggering an overall shift in AAA domains, and ultimately an iris-like contraction of the AAA ring.
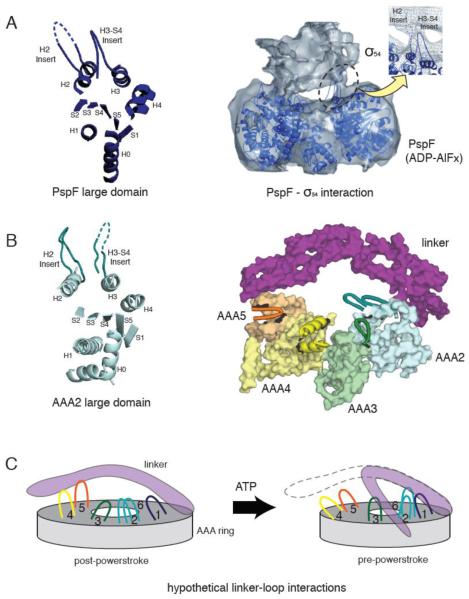
(A) The crystal structure of a PspF large domain showing the position of the H2 and H3-S4 insert loops (left). A 20 Å resolution cryo-electron microscopy reconstruction of the PspF hexamer in complex with its binding partner, σ54 in the presence of ADP-AlFx (right). The PspF hexamer crystal structure is docked into the electron microscopy structure, showing the H2 and H3-S4 insert loops at the site of PspF - σ54 interaction (reprinted from [46] with permission). (B) The crystal structure of AAA2 large domain in yeast cytoplasmic dynein, showing a similar position of the H2 and H3-S4 insert loops compared with PspF (left). The position of the unique loop inserts in the large domains of dynein AAA2-5, with respect to the linker (right). AAA1 and AAA6 are not shown for clarity. Insert loops lying at the top surface of the ring are depicted in cartoon format. The loops provide potential docking sites for the linker or dynein regulatory proteins. (C) A speculative model of how the loop insertions could act as docking sites for the linker at different stages of the ATPase cycle.
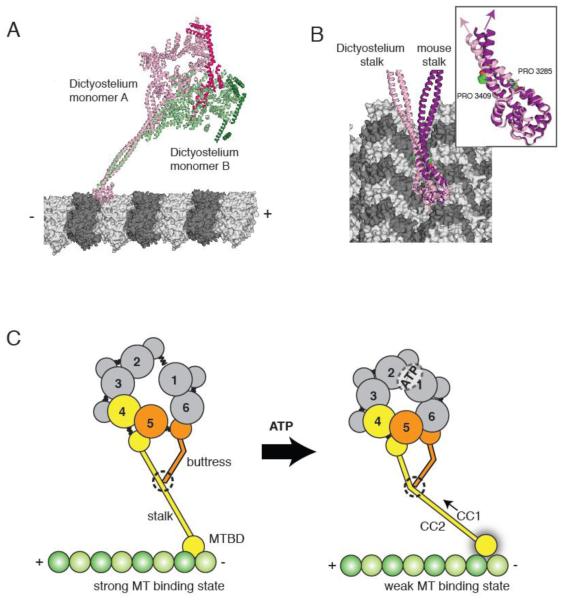
(A) The two Dictyostelium dynein monomers (monomer A in pink monomer B in green; linkers highlighted in darker hues) modeled onto microtubules based on docking of the mouse microtubule binding domain as in [30]. The different stalk angles of monomer A and B results in a different orientation of the dynein heads with respect to the microtubule longitudinal axis. (B) The stalks of the Dictyostelium stalk (pink) and mouse stalk (purple) viewed from the microtubule minus end. The inset shows the difference in stalk angle, which might be due to differences in the vicinity of the proline kinks in CC1 and CC2. (C) A model for stalk communication. Upon ATP binding, movements of AAA4 and AAA5 small domains are relayed to the stalk and buttress coiled coil extensions, respectively. Due to its interaction between the stalk, the buttress can push or pull on CC1, causing relative sliding motions between CC1 and CC2 and ultimately changing the microtubule binding affinity at the tip of the stalk.
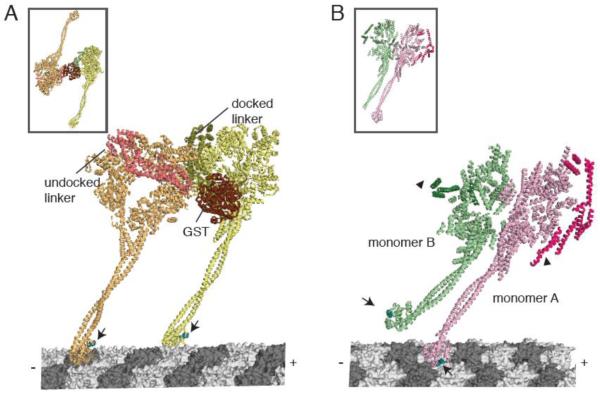
(A) A model for dimeric yeast dynein docked onto microtubules. (Inset shows the dimeric conformation in the crystal structure). The front head (orange) has an undocked linker (salmon) and a small kink in the stalk to accommodate a docked microtubule binding domain conformation. The rear head (yellow) has a docked linker and a straight stalk. The linkers are connected at the N-terminus by a GST dimerization domain. Arrows indicate equivalent positions in the microtubule binding domain (cyan). The linker face of the rear head is facing the C-terminal face of the front head. (B) The Dictyostelium dynein crystal structure (inset), showing the stacked conformation of monomer A (pink) and B (green). Using this crystal structure, monomer A was docked onto the microtubule. The putative conformation of the microtubule binding domain in monomer B was modeled based on the conformation of the distal stalk in monomer A. Arrows indicate equivalent positions in the microtubule binding domain (cyan), which are pointing in different directions. Arrowheads indicate the N-terminus of the linkers that must connect to a dimerization domain. In this configuration, the linkers are on the outside and the C-termini are on the inside of the stacked AAA rings.
References
-
- Gibbons IR, Rowe AJ. Dynein: A Protein with Adenosine Triphosphatase Activity from Cilia. Science. 1965;149:424–426. - PubMed
-
- Vallee RB, Williams JC, Varma D, Barnhart LE. Dynein: An ancient motor protein involved in multiple modes of transport. J Neurobiol. 2004;58:189–200. - PubMed
-
- Neuwald AF, Aravind L, Spouge JL, Koonin EV. AAA+: A class of chaperone-like ATPases associated with the assembly, operation, and disassembly of protein complexes. Genome Res. 1999;9:27–43. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
