

Nucleoid-enriched proteomes in developing plastids and chloroplasts from maize leaves: a new conceptual framework for nucleoid functions
- PMID: 22065420
- PMCID: PMC3252073
- DOI: 10.1104/pp.111.188474
Nucleoid-enriched proteomes in developing plastids and chloroplasts from maize leaves: a new conceptual framework for nucleoid functions
Abstract
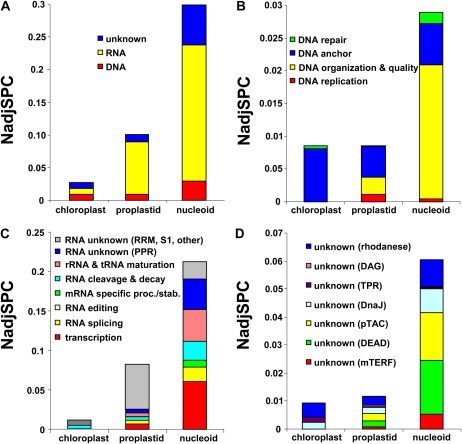
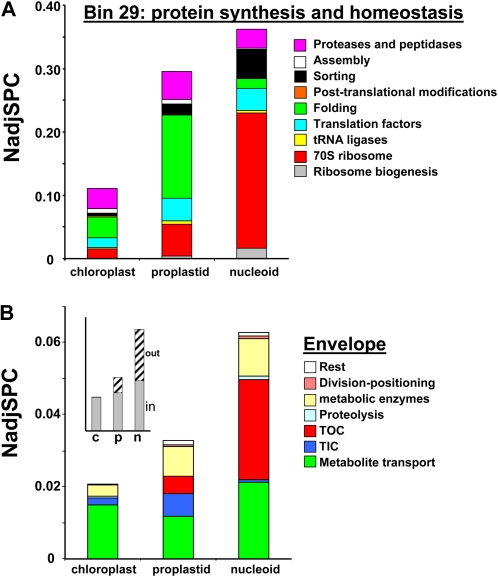
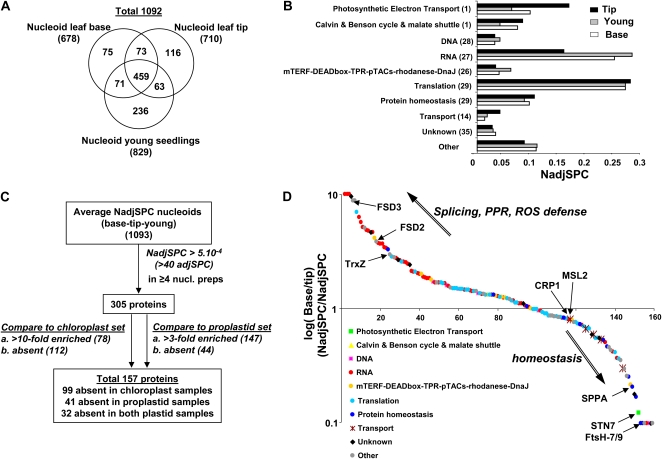
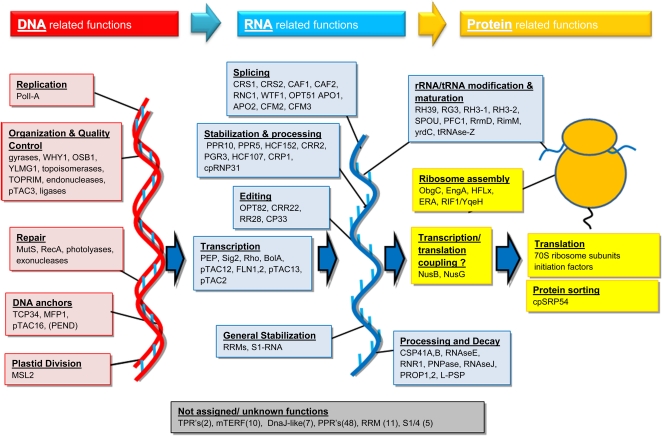
Plastids contain multiple copies of the plastid chromosome, folded together with proteins and RNA into nucleoids. The degree to which components of the plastid gene expression and protein biogenesis machineries are nucleoid associated, and the factors involved in plastid DNA organization, repair, and replication, are poorly understood. To provide a conceptual framework for nucleoid function, we characterized the proteomes of highly enriched nucleoid fractions of proplastids and mature chloroplasts isolated from the maize (Zea mays) leaf base and tip, respectively, using mass spectrometry. Quantitative comparisons with proteomes of unfractionated proplastids and chloroplasts facilitated the determination of nucleoid-enriched proteins. This nucleoid-enriched proteome included proteins involved in DNA replication, organization, and repair as well as transcription, mRNA processing, splicing, and editing. Many proteins of unknown function, including pentatricopeptide repeat (PPR), tetratricopeptide repeat (TPR), DnaJ, and mitochondrial transcription factor (mTERF) domain proteins, were identified. Strikingly, 70S ribosome and ribosome assembly factors were strongly overrepresented in nucleoid fractions, but protein chaperones were not. Our analysis strongly suggests that mRNA processing, splicing, and editing, as well as ribosome assembly, take place in association with the nucleoid, suggesting that these processes occur cotranscriptionally. The plastid developmental state did not dramatically change the nucleoid-enriched proteome but did quantitatively shift the predominating function from RNA metabolism in undeveloped plastids to translation and homeostasis in chloroplasts. This study extends the known maize plastid proteome by hundreds of proteins, including more than 40 PPR and mTERF domain proteins, and provides a resource for targeted studies on plastid gene expression. Details of protein identification and annotation are provided in the Plant Proteome Database.
Figures



Similar articles
-
A chloroplast-localized PPR protein required for plastid ribosome accumulation.Plant J. 2003 Dec;36(5):675-86. doi: 10.1046/j.1365-313x.2003.01915.x. Plant J. 2003. PMID: 14617068
-
Multifunctionality of plastid nucleoids as revealed by proteome analyses.Biochim Biophys Acta. 2016 Aug;1864(8):1016-38. doi: 10.1016/j.bbapap.2016.03.009. Epub 2016 Mar 14. Biochim Biophys Acta. 2016. PMID: 26987276 Review.
-
Evolution of RLSB, a nuclear-encoded S1 domain RNA binding protein associated with post-transcriptional regulation of plastid-encoded rbcL mRNA in vascular plants.BMC Evol Biol. 2016 Jun 29;16(1):141. doi: 10.1186/s12862-016-0713-1. BMC Evol Biol. 2016. PMID: 27356975 Free PMC article.
-
Emb15 encodes a plastid ribosomal assembly factor essential for embryogenesis in maize.Plant J. 2021 Apr;106(1):214-227. doi: 10.1111/tpj.15160. Epub 2021 Feb 11. Plant J. 2021. PMID: 33450100
-
Plastid Nucleoids: Insights into Their Shape and Dynamics.Plant Cell Physiol. 2024 May 14;65(4):551-559. doi: 10.1093/pcp/pcad090. Plant Cell Physiol. 2024. PMID: 37542434 Review.
Cited by
-
Comparative proteomics of chloroplasts envelopes from bundle sheath and mesophyll chloroplasts reveals novel membrane proteins with a possible role in c4-related metabolite fluxes and development.Front Plant Sci. 2013 Mar 28;4:65. doi: 10.3389/fpls.2013.00065. eCollection 2013. Front Plant Sci. 2013. PMID: 23543921 Free PMC article.
-
Dual targeting and retrograde translocation: regulators of plant nuclear gene expression can be sequestered by plastids.Int J Mol Sci. 2012;13(9):11085-11101. doi: 10.3390/ijms130911085. Epub 2012 Sep 6. Int J Mol Sci. 2012. PMID: 23109840 Free PMC article. Review.
-
Oleic acid-dependent modulation of NITRIC OXIDE ASSOCIATED1 protein levels regulates nitric oxide-mediated defense signaling in Arabidopsis.Plant Cell. 2012 Apr;24(4):1654-74. doi: 10.1105/tpc.112.096768. Epub 2012 Apr 6. Plant Cell. 2012. PMID: 22492810 Free PMC article.
-
WHIRLIES Are Multifunctional DNA-Binding Proteins With Impact on Plant Development and Stress Resistance.Front Plant Sci. 2022 Apr 21;13:880423. doi: 10.3389/fpls.2022.880423. eCollection 2022. Front Plant Sci. 2022. PMID: 35528945 Free PMC article. Review.
-
Seedling Lethal1, a pentatricopeptide repeat protein lacking an E/E+ or DYW domain in Arabidopsis, is involved in plastid gene expression and early chloroplast development.Plant Physiol. 2013 Dec;163(4):1844-58. doi: 10.1104/pp.113.227199. Epub 2013 Oct 21. Plant Physiol. 2013. PMID: 24144791 Free PMC article.
References
-
- Albrecht V, Ingenfeld A, Apel K. (2008) Snowy cotyledon 2: the identification of a zinc finger domain protein essential for chloroplast development in cotyledons but not in true leaves. Plant Mol Biol 66: 599–608 - PubMed
-
- Aravind L, Koonin EV. (2000) SAP: a putative DNA-binding motif involved in chromosomal organization. Trends Biochem Sci 25: 112–114 - PubMed
-
- Arsova B, Hoja U, Wimmelbacher M, Greiner E, Ustun S, Melzer M, Petersen K, Lein W, Bornke F. (2010) Plastidial thioredoxin z interacts with two fructokinase-like proteins in a thiol-dependent manner: evidence for an essential role in chloroplast development in Arabidopsis and Nicotiana benthamiana. Plant Cell 22: 1498–1515 - PMC - PubMed
-
- Babiychuk E, Vandepoele K, Wissing J, Garcia-Diaz M, De Rycke R, Akbari H, Joubes J, Beeckman T, Jansch L, Frentzen M, et al. (2011) Plastid gene expression and plant development require a plastidic protein of the mitochondrial transcription termination factor family. Proc Natl Acad Sci USA 108: 6674–6679 - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
