

Synthetic double-stranded RNA induces innate immune responses similar to a live viral vaccine in humans
- PMID: 22065672
- PMCID: PMC3256967
- DOI: 10.1084/jem.20111171
Synthetic double-stranded RNA induces innate immune responses similar to a live viral vaccine in humans
Abstract
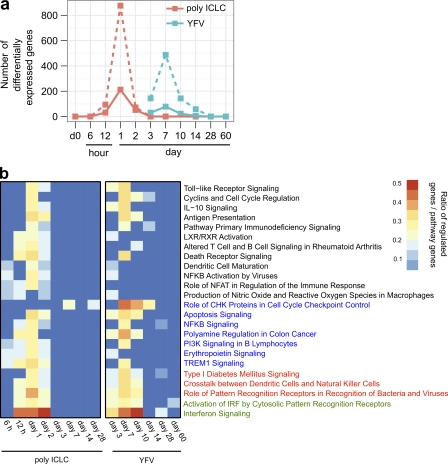
Adjuvants are critical for the success of vaccines. Agonists of microbial pattern recognition receptors (PRRs) are promising new adjuvant candidates. A mechanism through which adjuvants enhance immune responses is to stimulate innate immunity. We studied the innate immune response in humans to synthetic double-stranded RNA (polyinosinic:polycytidylic acid [poly IC] stabilized with poly-L-lysine [poly ICLC]), an agonist for toll-like receptor (TLR) 3, and the cytosolic RNA helicase MDA-5. Transcriptional analysis of blood samples from eight volunteers, after subcutaneous administration of poly ICLC, showed up-regulation of genes involved in multiple innate immune pathways in all subjects, including interferon (IFN) and inflammasome signaling. Blocking type I IFN receptor ex vivo significantly dampened the response to poly IC. Comparative transcriptional analysis showed that several innate immune pathways were similarly induced in volunteers immunized with the highly efficacious yellow fever vaccine. Therefore, a chemically defined PRR agonist like poly ICLC can be a reliable and authentic microbial mimic for inducing innate immune responses in humans.
Figures
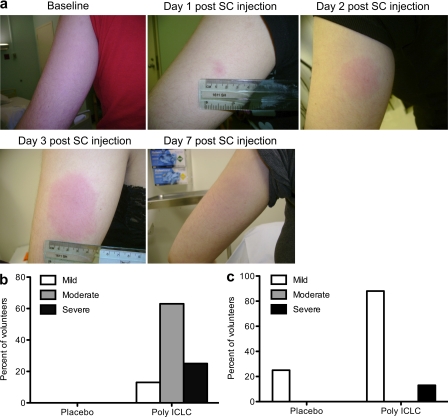
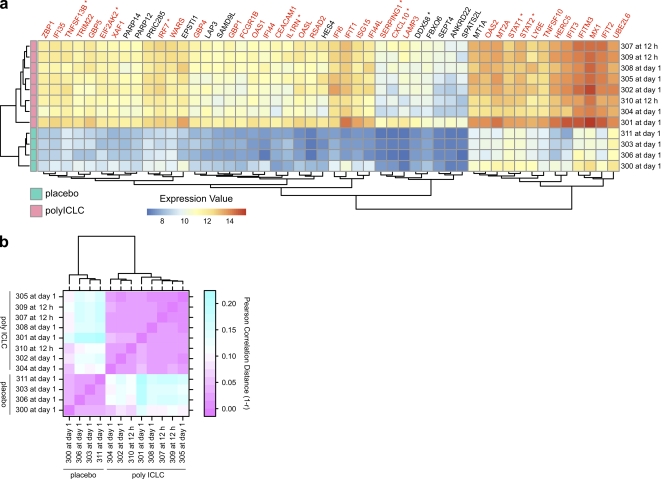
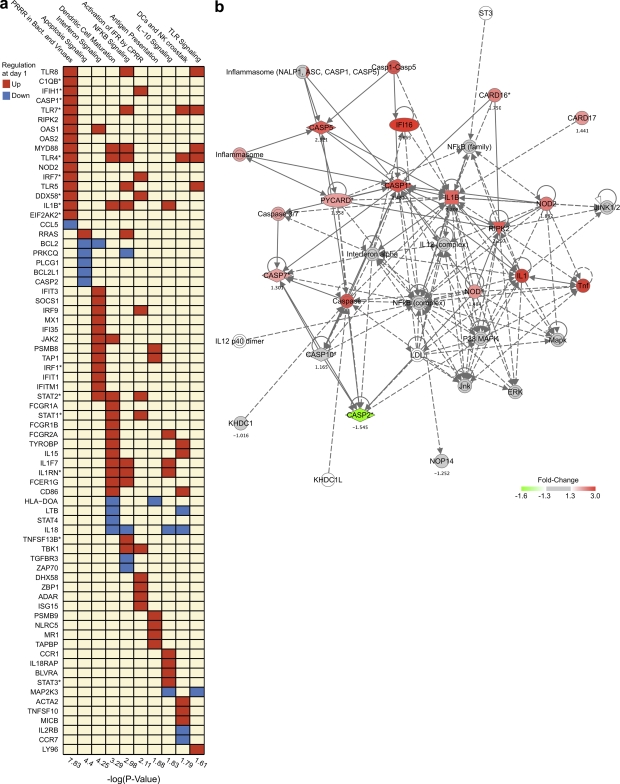
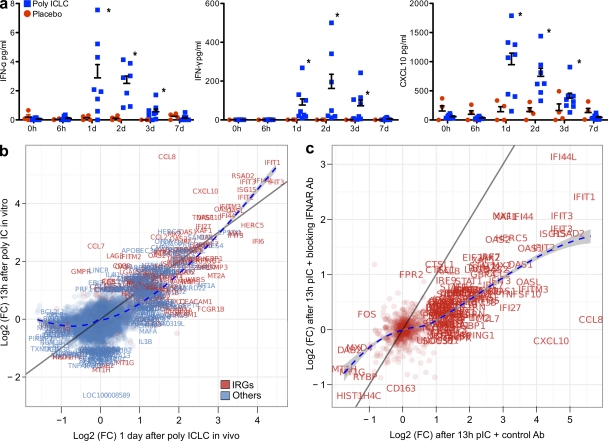
Comment in
-
Learning vaccinology from viral infections.J Exp Med. 2011 Nov 21;208(12):2347-9. doi: 10.1084/jem.20112321. J Exp Med. 2011. PMID: 22110181 Free PMC article.
References
-
- Benjamini Y., Hochberg Y. 1995. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. R. Stat. Soc. Series B Stat. Methodol. 57:289–300
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
