

Role of type 2 deiodinase in response to acute lung injury (ALI) in mice
- PMID: 22065740
- PMCID: PMC3241808
- DOI: 10.1073/pnas.1109926108
Role of type 2 deiodinase in response to acute lung injury (ALI) in mice
Abstract
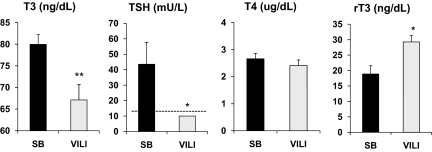
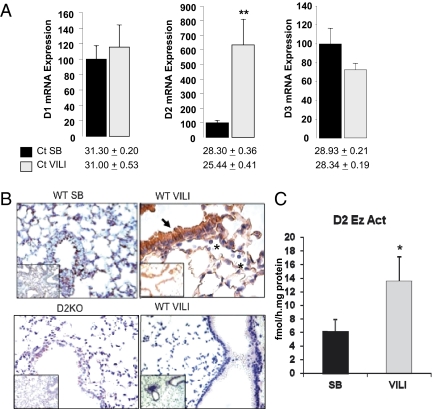
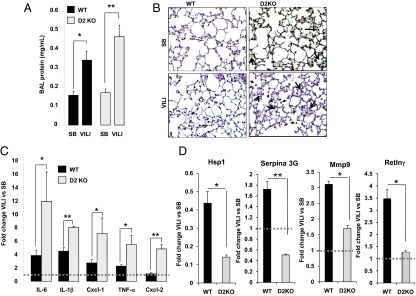
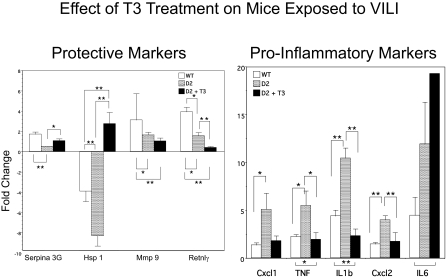
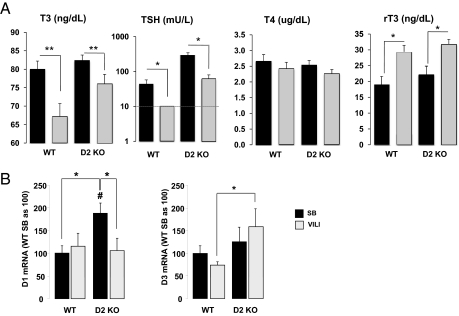
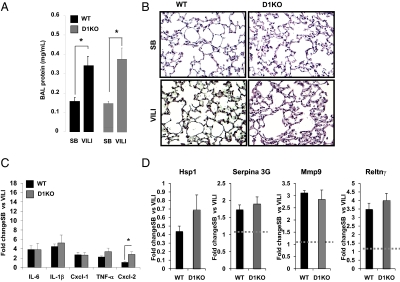
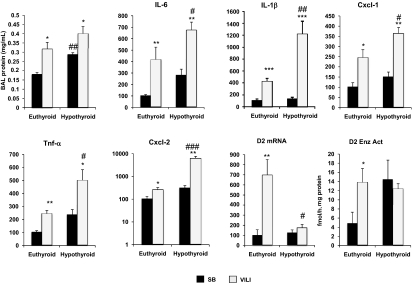
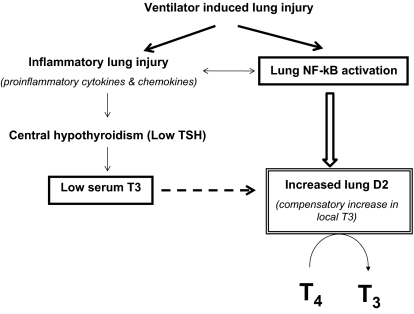
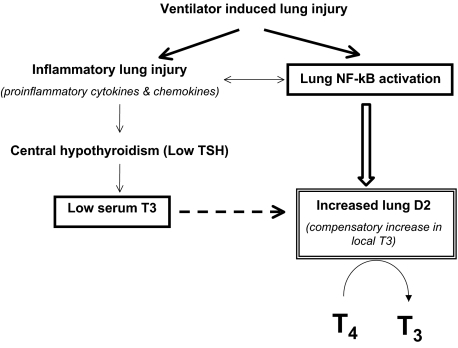
Thyroid hormone (TH) metabolism, mediated by deiodinase types 1, 2, and 3 (D1, D2, and D3) is profoundly affected by acute illness. We examined the role of TH metabolism during ventilator-induced lung injury (VILI) in mice. Mice exposed to VILI recapitulated the serum TH findings of acute illness, namely a decrease in 3,5,3'-triiodothyronine (T(3)) and thyroid-stimulating hormone and an increase in reverse T(3). Both D2 immunoreactivity and D2 enzymatic activity were increased significantly. D1 and D3 activity did not change. Using D2 knockout (D2KO) mice, we determined whether the increase in D2 was an adaptive response. Although similar changes in serum TH levels were observed in D2KO and WT mice, D2KO mice exhibited greater susceptibility to VILI than WT mice, as evidenced by poorer alveoli integrity and quantified by lung chemokine and cytokine mRNA induction. These data suggest that an increase in lung D2 is protective against VILI. Similar findings of increased inflammatory markers were found in hypothyroid WT mice exposed to VILI compared with euthyroid mice, indicating that the lungs were functionally hypothyroid. Treatment of D2KO mice with T(3) reversed many of the lung chemokine and cytokine profiles seen in response to VILI, demonstrating a role for T(3) in the treatment of lung injury. We conclude that TH metabolism in the lung is linked to the response to inflammatory injury and speculate that D2 exerts its protective effect by making more TH available to the injured lung tissue.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Bianco AC, Salvatore D, Gereben B, Berry MJ, Larsen PR. Biochemistry, cellular and molecular biology, and physiological roles of the iodothyronine selenodeiodinases. Endocr Rev. 2002;23(1):38–89. - PubMed
-
- Alkemade A. Central and peripheral effects of thyroid hormone signalling in the control of energy metabolism. J Neuroendocrinol. 2010;22(1):56–63. - PubMed
-
- Williams GR. Neurodevelopmental and neurophysiological actions of thyroid hormone. J Neuroendocrinol. 2008;20:784–794. - PubMed
-
- Gogakos AI, Duncan Bassett JH, Williams GR. Thyroid and bone. Arch Biochem Biophys. 2010;503(1):129–136. - PubMed
-
- Ness GC. Thyroid hormone. Basis for its hypocholesterolemic effect. J Fla Med Assoc. 1991;78:383–385. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
