

Batf3-dependent CD11b(low/-) peripheral dendritic cells are GM-CSF-independent and are not required for Th cell priming after subcutaneous immunization
- PMID: 22065991
- PMCID: PMC3196467
- DOI: 10.1371/journal.pone.0025660
Batf3-dependent CD11b(low/-) peripheral dendritic cells are GM-CSF-independent and are not required for Th cell priming after subcutaneous immunization
Abstract
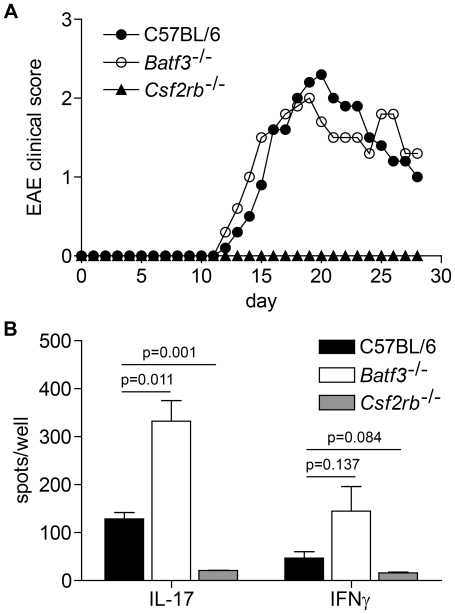
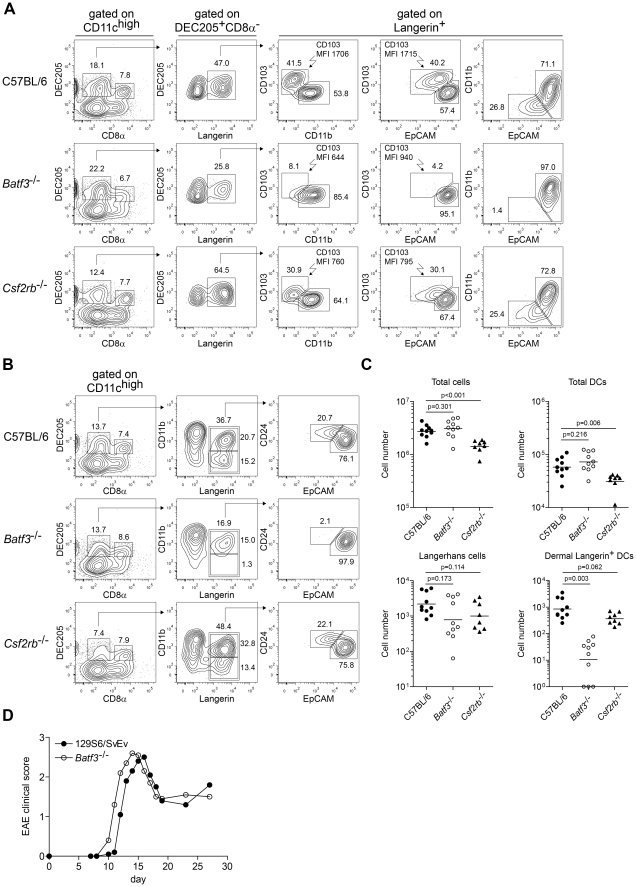
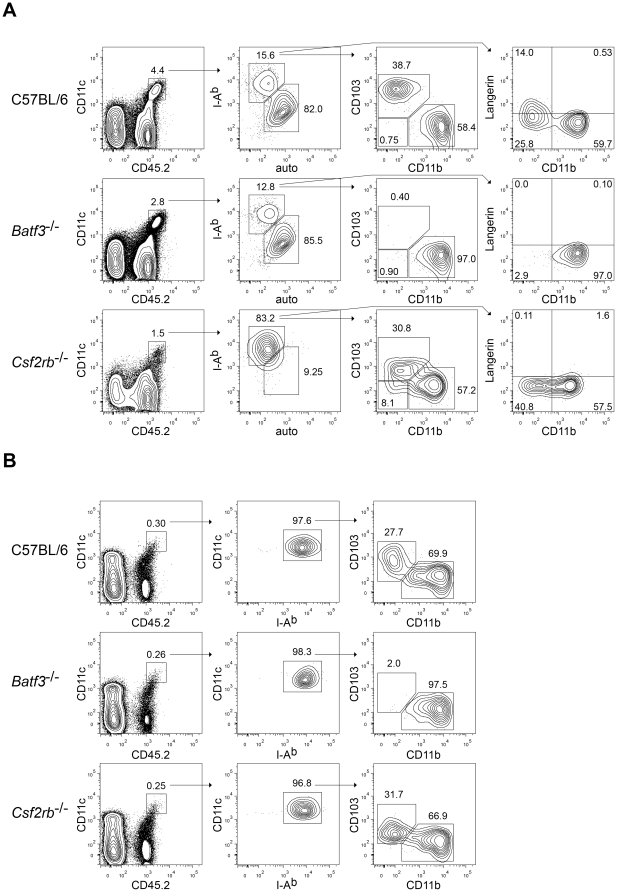
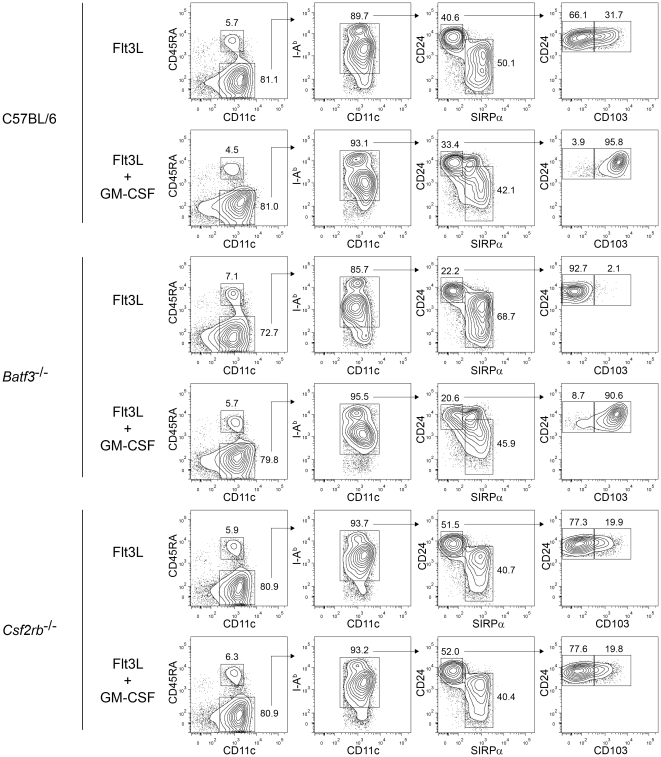
Dendritic cells (DCs) subsets differ in precursor cell of origin, functional properties, requirements for growth factors, and dependence on transcription factors. Lymphoid-tissue resident CD8α(+) conventional DCs (cDCs) and CD11b(low/-)CD103(+) non-lymphoid DCs are developmentally related, each being dependent on FMS-like tyrosine kinase 3 ligand (Flt3L), and requiring the transcription factors Batf3, Irf8, and Id2 for development. It was recently suggested that granulocyte/macrophage colony stimulating factor (GM-CSF) was required for the development of dermal CD11b(low/-)Langerin(+)CD103(+) DCs, and that this dermal DC subset was required for priming autoreactive T cells in experimental autoimmune encephalitis (EAE). Here, we compared development of peripheral tissue DCs and susceptibility to EAE in GM-CSF receptor deficient (Csf2rb(-/-)) and Batf3(-/-) mice. We find that Batf3-dependent dermal CD11b(low/-)Langerin(+) DCs do develop in Csf2rb(-/-) mice, but that they express reduced, but not absent, levels of CD103. Further, Batf3(-/-) mice lacking all peripheral CD11b(low/-) DCs show robust Th cell priming after subcutaneous immunization and are susceptible to EAE. Our results suggest that defective T effector priming and resistance to EAE exhibited by Csf2rb(-/-) mice does not result from the absence of dermal CD11b(low/-)Langerin(+)CD103(+) DCs.
Conflict of interest statement
Figures
References
-
- Hamilton JA. Colony-stimulating factors in inflammation and autoimmunity. Nat Rev Immunol. 2008;8:533–544. - PubMed
-
- Ponomarev ED, Shriver LP, Maresz K, Pedras-Vasconcelos J, Verthelyi D, et al. GM-CSF production by autoreactive T cells is required for the activation of microglial cells and the onset of experimental autoimmune encephalomyelitis. J Immunol. 2007;178:39–48. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
