

Calcium control of neurotransmitter release
- PMID: 22068972
- PMCID: PMC3249630
- DOI: 10.1101/cshperspect.a011353
Calcium control of neurotransmitter release
Abstract
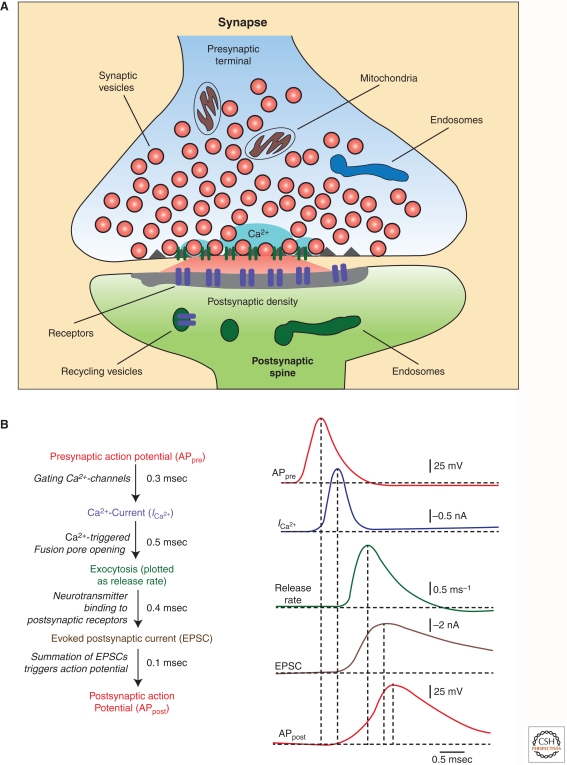
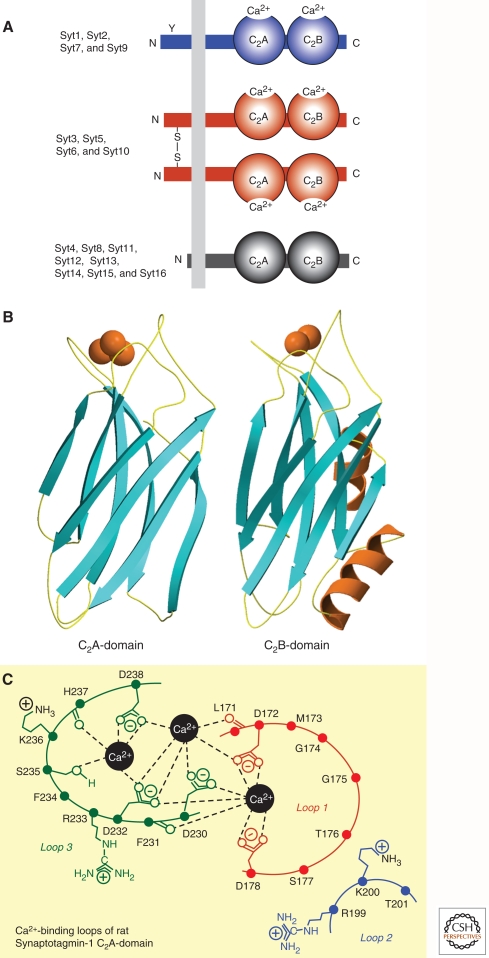
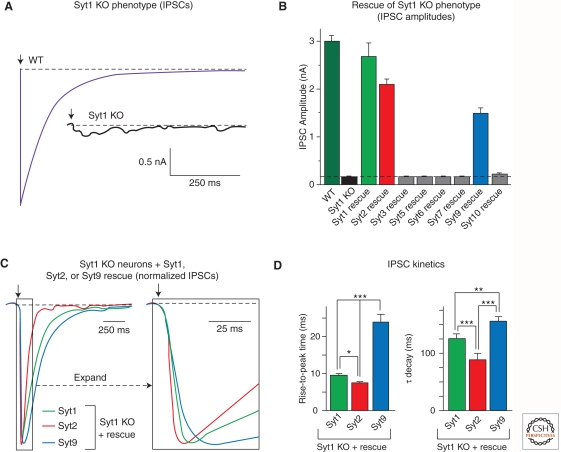
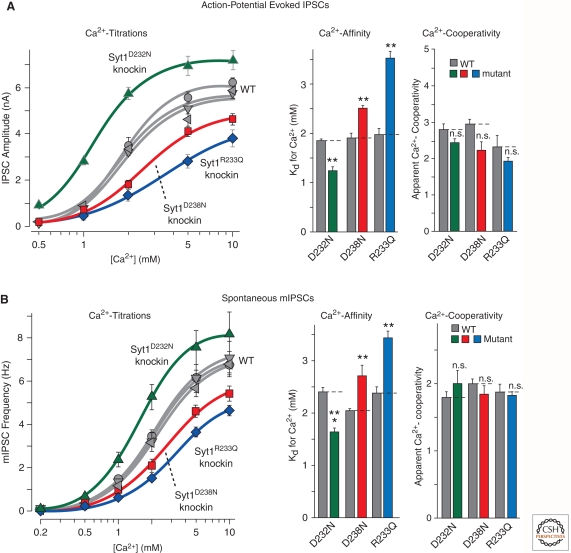
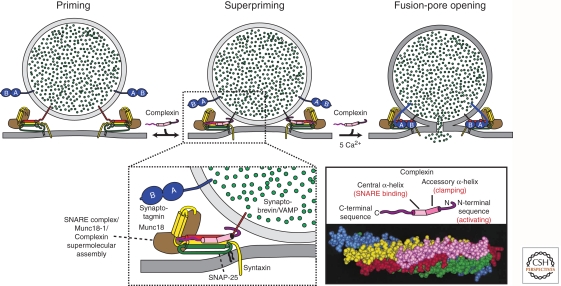
Upon entering a presynaptic terminal, an action potential opens Ca(2+) channels, and transiently increases the local Ca(2+) concentration at the presynaptic active zone. Ca(2+) then triggers neurotransmitter release within a few hundred microseconds by activating synaptotagmins Ca(2+). Synaptotagmins bind Ca(2+) via two C2-domains, and transduce the Ca(2+) signal into a nanomechanical activation of the membrane fusion machinery; this activation is mediated by the Ca(2+)-dependent interaction of the synaptotagmin C2-domains with phospholipids and SNARE proteins. In triggering exocytosis, synaptotagmins do not act alone, but require an obligatory cofactor called complexin, a small protein that binds to SNARE complexes and simultaneously activates and clamps the SNARE complexes, thereby positioning the SNARE complexes for subsequent synaptotagmin action. The conserved function of synaptotagmins and complexins operates generally in most, if not all, Ca(2+)-regulated forms of exocytosis throughout the body in addition to synaptic vesicle exocytosis, including in the degranulation of mast cells, acrosome exocytosis in sperm cells, hormone secretion from endocrine cells, and neuropeptide release.
Figures
References
-
- Andrews NW, Chakrabarti S 2005. There’s more to life than neurotransmission: The regulation of exocytosis by synaptotagmin VII. Trends Cell Biol 15: 626–631 - PubMed
-
- Bennett MK, Calakos N, Scheller RH 1992. Syntaxin: A synaptic protein implicated in docking of synaptic vesicles at presynaptic active zones. Science 257: 255–259 - PubMed
-
- Brose N, Petrenko AG, Südhof TC, Jahn R 1992. Synaptotagmin: A Ca2+ sensor on the synaptic vesicle surface. Science 256: 1021–1025 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous