

The astrocyte-expressed integrin αvβ8 governs blood vessel sprouting in the developing retina
- PMID: 22069187
- PMCID: PMC3253036
- DOI: 10.1242/dev.069153
The astrocyte-expressed integrin αvβ8 governs blood vessel sprouting in the developing retina
Abstract
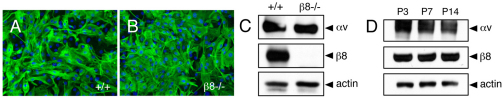
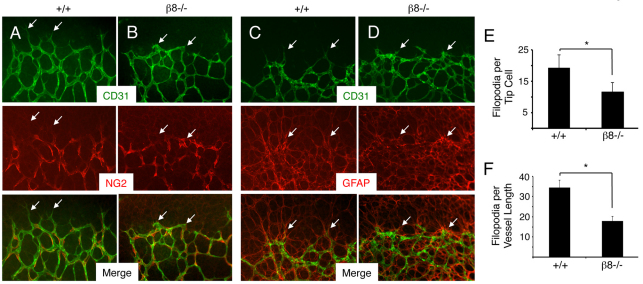
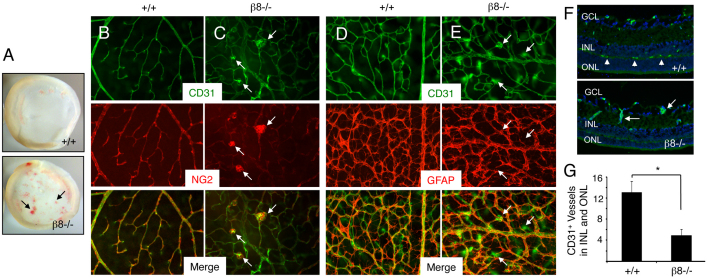
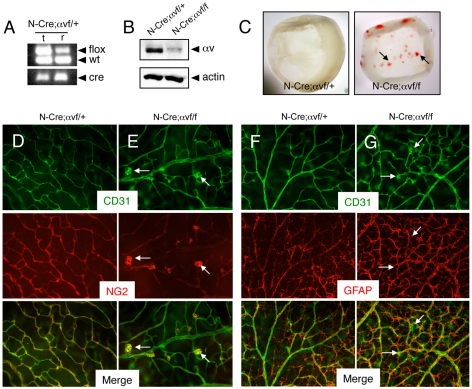
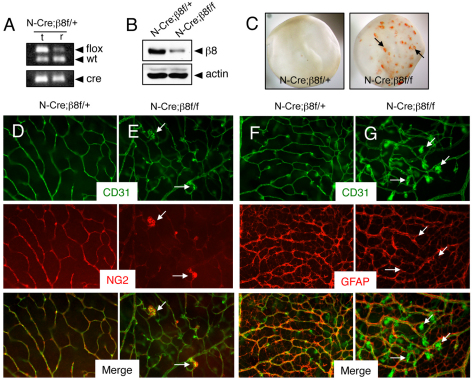
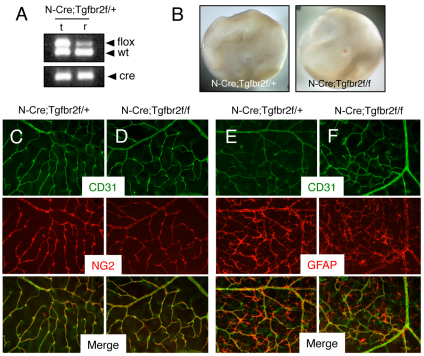
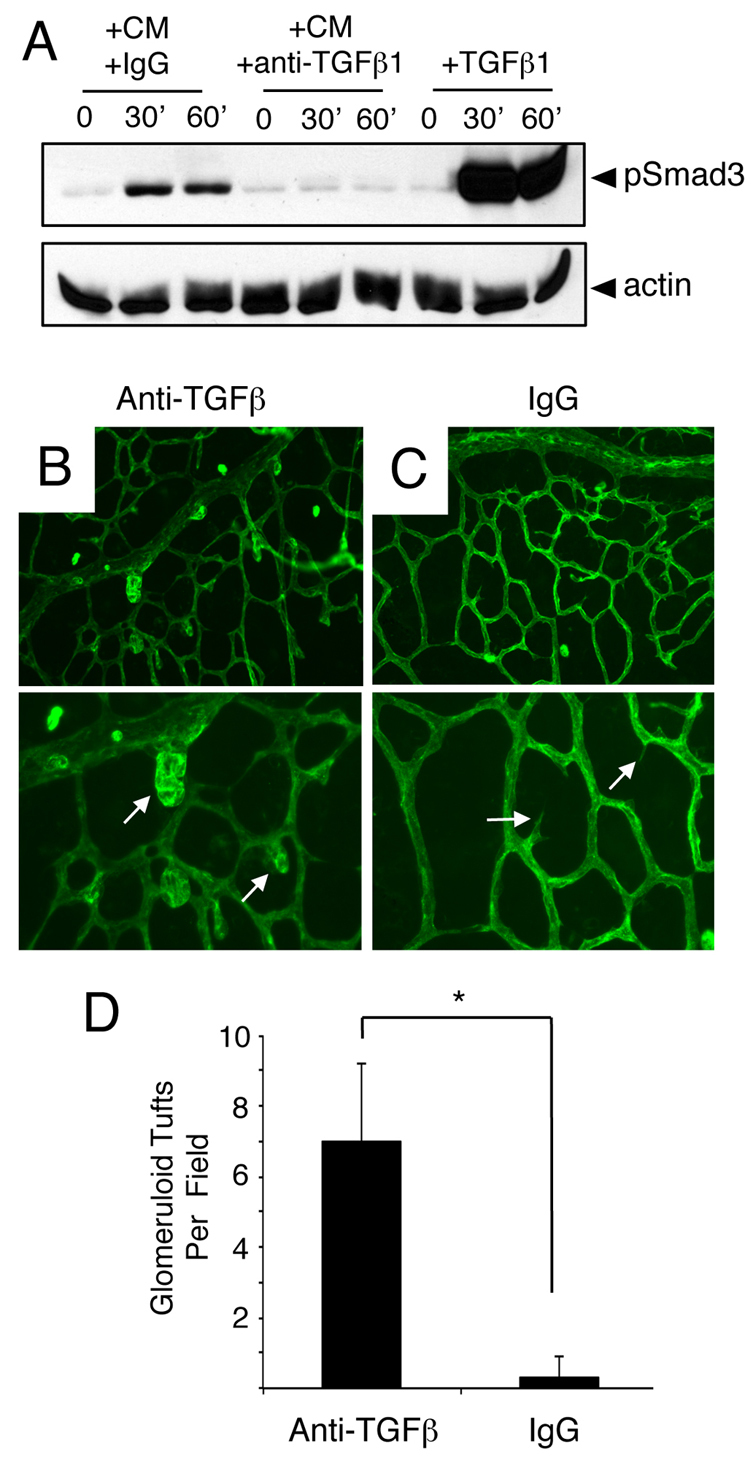
The mouse retina is vascularized after birth when angiogenic blood vessels grow and sprout along a pre-formed latticework of astrocytes. How astrocyte-derived cues control patterns of blood vessel growth and sprouting, however, remains enigmatic. Here, we have used molecular genetic strategies in mice to demonstrate that αvβ8 integrin expressed in astrocytes is essential for neovascularization of the developing retina. Selective ablation of αv or β8 integrin gene expression in astrocytes leads to impaired blood vessel sprouting and intraretinal hemorrhage, particularly during formation of the secondary vascular plexus. These pathologies correlate, in part, with diminished αvβ8 integrin-mediated activation of extracellular matrix-bound latent transforming growth factor βs (TGFβs) and defective TGFβ signaling in vascular endothelial cells, but not astrocytes. Collectively, our data demonstrate that αvβ8 integrin is a component of a paracrine signaling axis that links astrocytes to blood vessels and is essential for proper regulation of retinal angiogenesis.
Figures
References
-
- Benedito R., Roca C., Sorensen I., Adams S., Gossler A., Fruttiger M., Adams R. H. (2009). The notch ligands Dll4 and Jagged1 have opposing effects on angiogenesis. Cell 137, 1124–1135 - PubMed
-
- Benjamin L. E., Hemo I., Keshet E. (1998). A plasticity window for blood vessel remodelling is defined by pericyte coverage of the preformed endothelial network and is regulated by PDGF-B and VEGF. Development 125, 1591–1598 - PubMed
-
- Chen J., Smith L. E. (2007). Retinopathy of prematurity. Angiogenesis 10, 133–140 - PubMed
-
- Chytil A., Magnuson M. A., Wright C. V., Moses H. L. (2002). Conditional inactivation of the TGF-beta type II receptor using Cre:Lox. Genesis 32, 73–75 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
