

O-GlcNAcylation, novel post-translational modification linking myocardial metabolism and cardiomyocyte circadian clock
- PMID: 22069332
- PMCID: PMC3247942
- DOI: 10.1074/jbc.M111.278903
O-GlcNAcylation, novel post-translational modification linking myocardial metabolism and cardiomyocyte circadian clock
Abstract
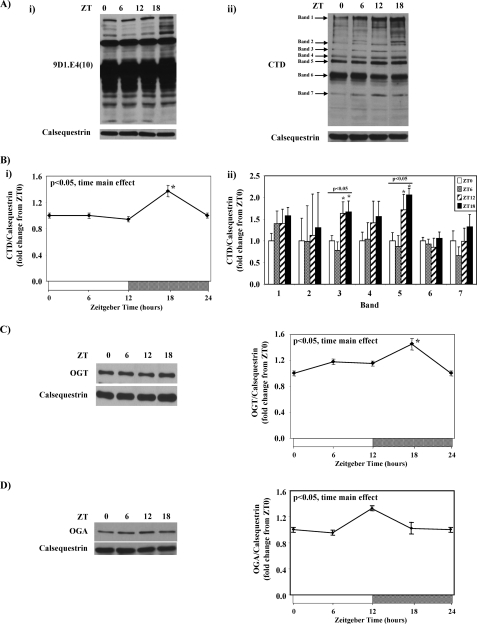
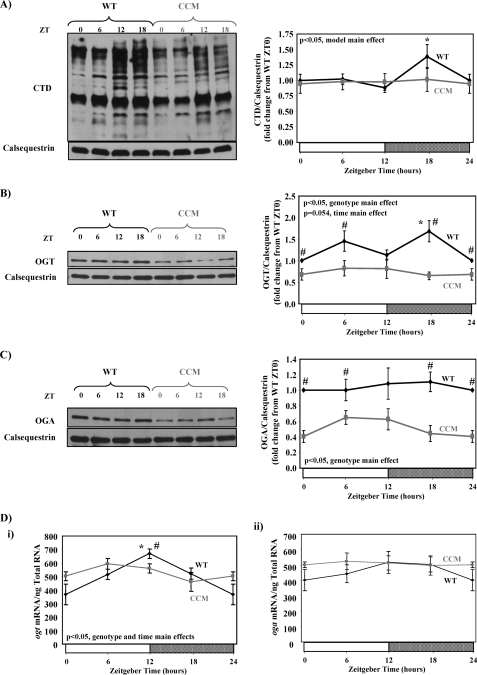
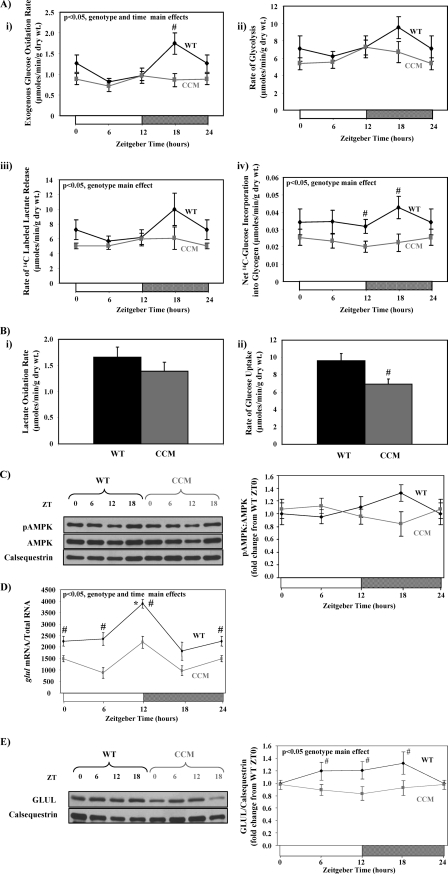
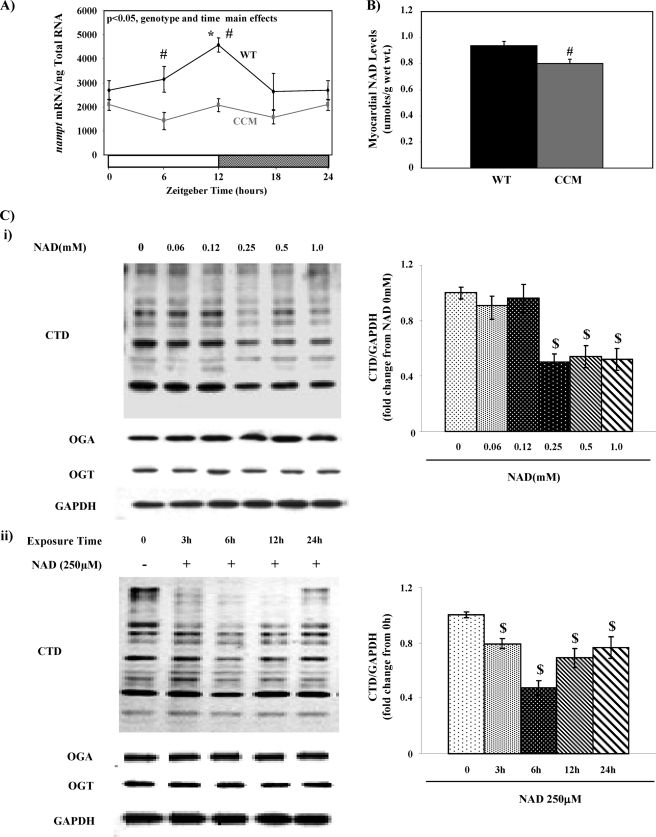
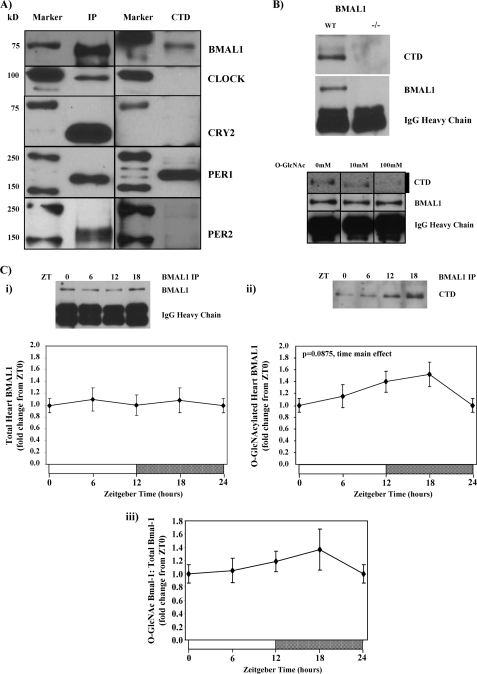
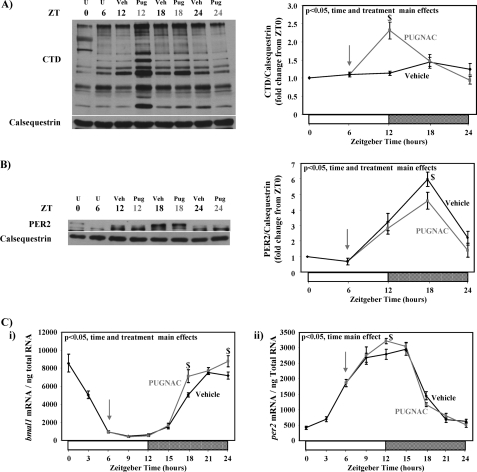
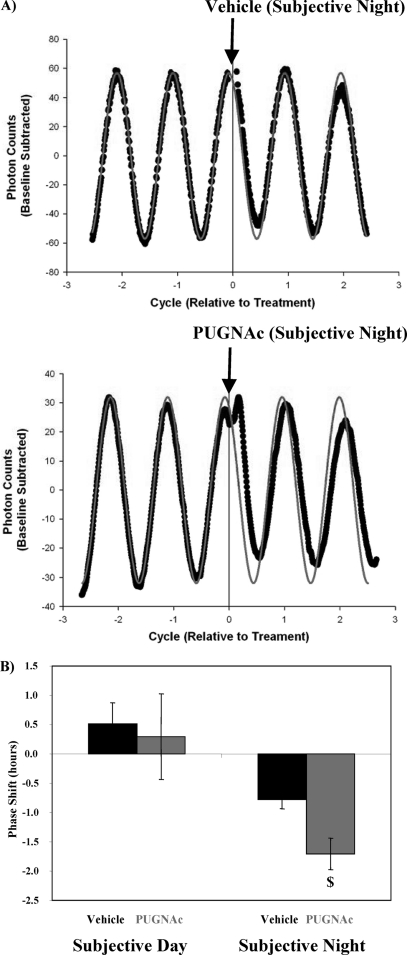
The cardiomyocyte circadian clock directly regulates multiple myocardial functions in a time-of-day-dependent manner, including gene expression, metabolism, contractility, and ischemic tolerance. These same biological processes are also directly influenced by modification of proteins by monosaccharides of O-linked β-N-acetylglucosamine (O-GlcNAc). Because the circadian clock and protein O-GlcNAcylation have common regulatory roles in the heart, we hypothesized that a relationship exists between the two. We report that total cardiac protein O-GlcNAc levels exhibit a diurnal variation in mouse hearts, peaking during the active/awake phase. Genetic ablation of the circadian clock specifically in cardiomyocytes in vivo abolishes diurnal variations in cardiac O-GlcNAc levels. These time-of-day-dependent variations appear to be mediated by clock-dependent regulation of O-GlcNAc transferase and O-GlcNAcase protein levels, glucose metabolism/uptake, and glutamine synthesis in an NAD-independent manner. We also identify the clock component Bmal1 as an O-GlcNAc-modified protein. Increasing protein O-GlcNAcylation (through pharmacological inhibition of O-GlcNAcase) results in diminished Per2 protein levels, time-of-day-dependent induction of bmal1 gene expression, and phase advances in the suprachiasmatic nucleus clock. Collectively, these data suggest that the cardiomyocyte circadian clock increases protein O-GlcNAcylation in the heart during the active/awake phase through coordinated regulation of the hexosamine biosynthetic pathway and that protein O-GlcNAcylation in turn influences the timing of the circadian clock.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
