

The peripheral nervous system supports blood cell homing and survival in the Drosophila larva
- PMID: 22071105
- PMCID: PMC3222213
- DOI: 10.1242/dev.067322
The peripheral nervous system supports blood cell homing and survival in the Drosophila larva
Abstract
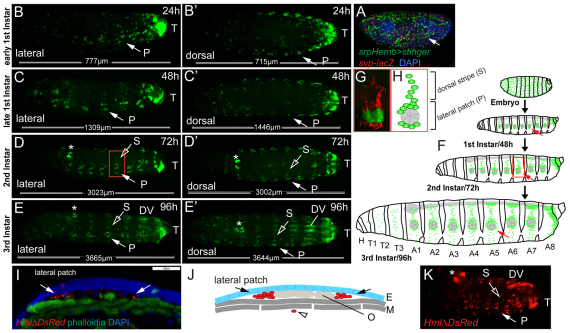
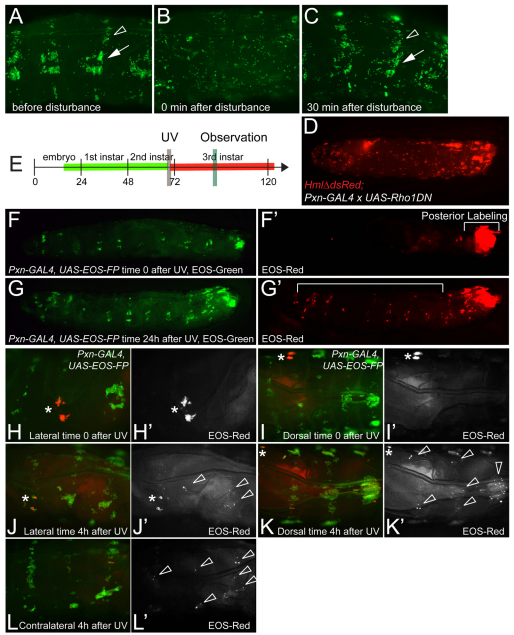
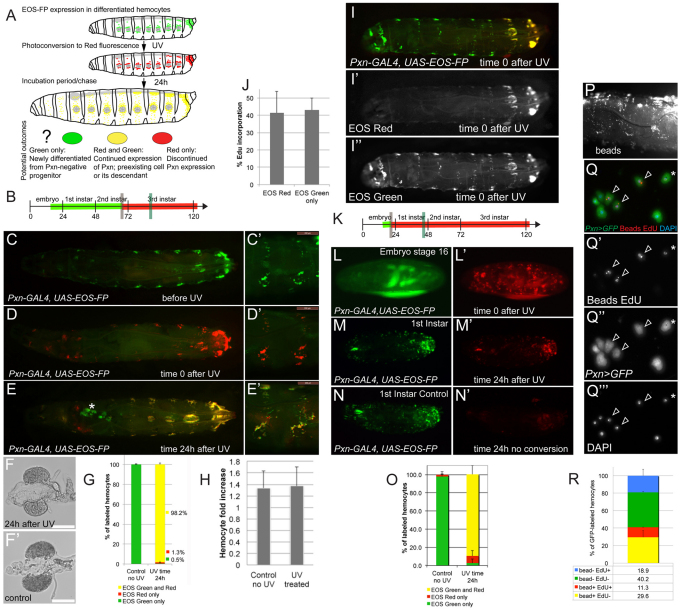
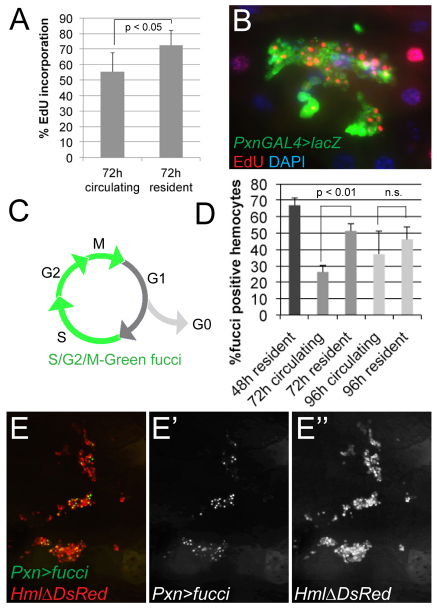
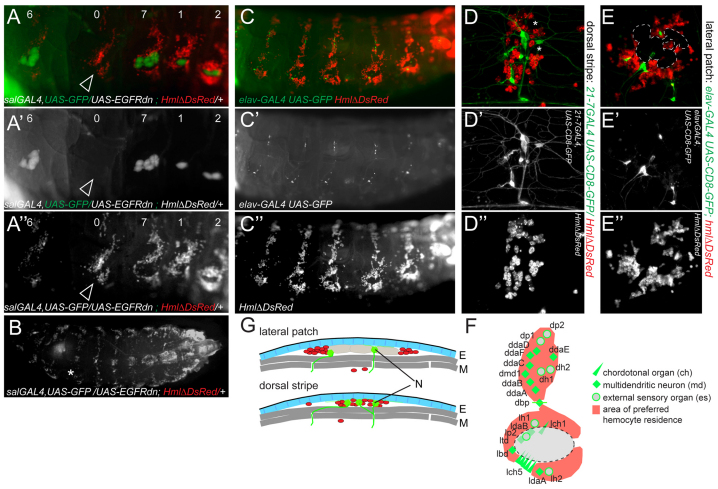
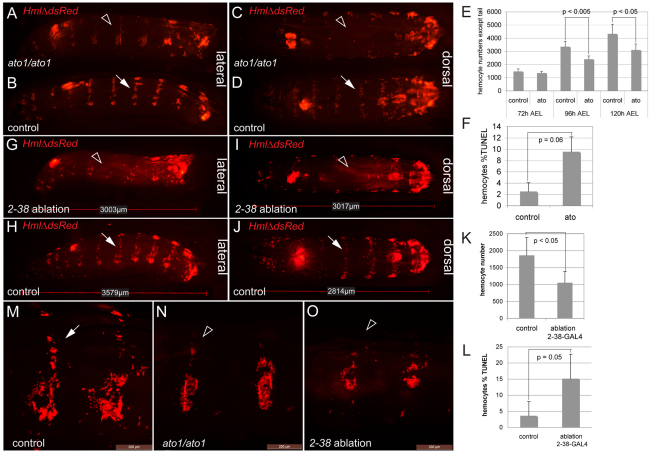
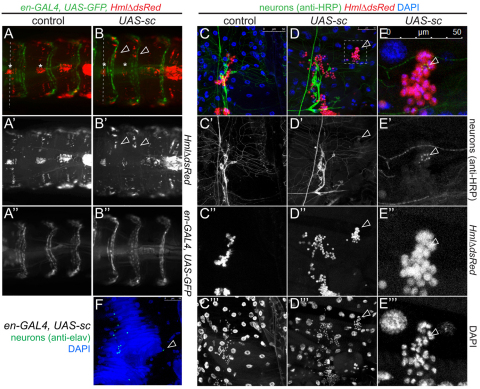
Interactions of hematopoietic cells with their microenvironment control blood cell colonization, homing and hematopoiesis. Here, we introduce larval hematopoiesis as the first Drosophila model for hematopoietic colonization and the role of the peripheral nervous system (PNS) as a microenvironment in hematopoiesis. The Drosophila larval hematopoietic system is founded by differentiated hemocytes of the embryo, which colonize segmentally repeated epidermal-muscular pockets and proliferate in these locations. Importantly, we show that these resident hemocytes tightly colocalize with peripheral neurons and we demonstrate that larval hemocytes depend on the PNS as an attractive and trophic microenvironment. atonal (ato) mutant or genetically ablated larvae, which are deficient for subsets of peripheral neurons, show a progressive apoptotic decline in hemocytes and an incomplete resident hemocyte pattern, whereas supernumerary peripheral neurons induced by ectopic expression of the proneural gene scute (sc) misdirect hemocytes to these ectopic locations. This PNS-hematopoietic connection in Drosophila parallels the emerging role of the PNS in hematopoiesis and immune functions in vertebrates, and provides the basis for the systematic genetic dissection of the PNS-hematopoietic axis in the future.
Figures
Similar articles
-
Of blood cells and the nervous system: hematopoiesis in the Drosophila larva.Fly (Austin). 2012 Oct-Dec;6(4):254-60. doi: 10.4161/fly.22267. Epub 2012 Sep 28. Fly (Austin). 2012. PMID: 23022764 Free PMC article. Review.
-
Regulation of Drosophila hematopoietic sites by Activin-β from active sensory neurons.Nat Commun. 2017 Jul 27;8:15990. doi: 10.1038/ncomms15990. Nat Commun. 2017. PMID: 28748922 Free PMC article.
-
Size control of the Drosophila hematopoietic niche by bone morphogenetic protein signaling reveals parallels with mammals.Proc Natl Acad Sci U S A. 2012 Feb 28;109(9):3389-94. doi: 10.1073/pnas.1109407109. Epub 2012 Feb 13. Proc Natl Acad Sci U S A. 2012. PMID: 22331866 Free PMC article.
-
Regulation of larval hematopoiesis in Drosophila melanogaster: a role for the multi sex combs gene.Genetics. 2002 Nov;162(3):1259-74. doi: 10.1093/genetics/162.3.1259. Genetics. 2002. PMID: 12454071 Free PMC article.
-
Drosophila as a Model to Study Cellular Communication Between the Hematopoietic Niche and Blood Progenitors Under Homeostatic Conditions and in Response to an Immune Stress.Front Immunol. 2021 Aug 16;12:719349. doi: 10.3389/fimmu.2021.719349. eCollection 2021. Front Immunol. 2021. PMID: 34484226 Free PMC article. Review.
Cited by
-
Schnurri regulates hemocyte function to promote tissue recovery after DNA damage.J Cell Sci. 2012 Mar 15;125(Pt 6):1393-400. doi: 10.1242/jcs.095323. Epub 2012 Jan 24. J Cell Sci. 2012. PMID: 22275438 Free PMC article.
-
Regulation of Drosophila Hematopoiesis in Lymph Gland: From a Developmental Signaling Point of View.Int J Mol Sci. 2020 Jul 24;21(15):5246. doi: 10.3390/ijms21155246. Int J Mol Sci. 2020. PMID: 32722007 Free PMC article. Review.
-
Cellular Pathogenesis of Chemotherapy-Induced Peripheral Neuropathy: Insights From Drosophila and Human-Engineered Skin Models.Front Pain Res (Lausanne). 2022 Jul 8;3:912977. doi: 10.3389/fpain.2022.912977. eCollection 2022. Front Pain Res (Lausanne). 2022. PMID: 35875478 Free PMC article. Review.
-
Fos regulates macrophage infiltration against surrounding tissue resistance by a cortical actin-based mechanism in Drosophila.PLoS Biol. 2022 Jan 6;20(1):e3001494. doi: 10.1371/journal.pbio.3001494. eCollection 2022 Jan. PLoS Biol. 2022. PMID: 34990456 Free PMC article.
-
Conserved chamber-specific polyploidy maintains heart function in Drosophila.Development. 2023 Aug 15;150(16):dev201896. doi: 10.1242/dev.201896. Epub 2023 Aug 22. Development. 2023. PMID: 37526609 Free PMC article.
References
-
- Adams G. B., Scadden D. T. (2006). The hematopoietic stem cell in its place. Nat. Immunol. 7, 333–337 - PubMed
-
- Ashburner M., Golic K. M., Hawley R. S. (2004). Drosophila: A Laboratory Handbook. New York: Cold Spring Harbor Laboratory Press;
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
