

Genetic correlations and the evolution of photoperiodic time measurement within a local population of the pitcher-plant mosquito, Wyeomyia smithii
- PMID: 22072069
- PMCID: PMC3330697
- DOI: 10.1038/hdy.2011.108
Genetic correlations and the evolution of photoperiodic time measurement within a local population of the pitcher-plant mosquito, Wyeomyia smithii
Abstract
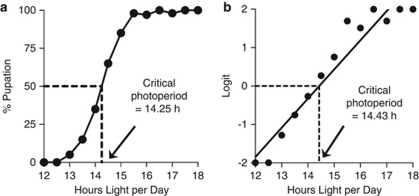
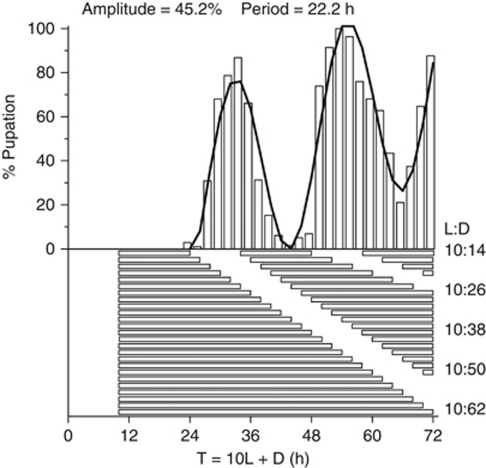
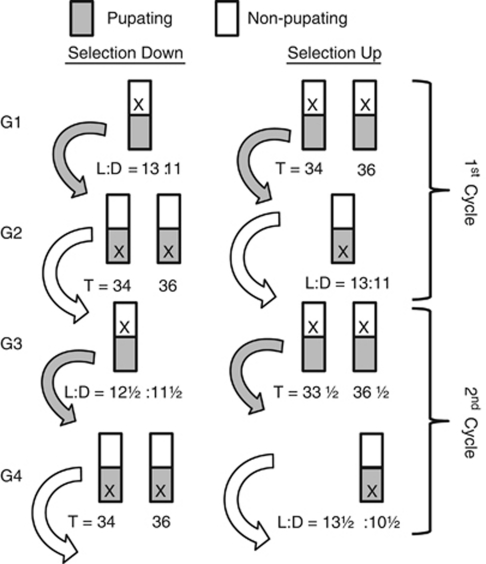
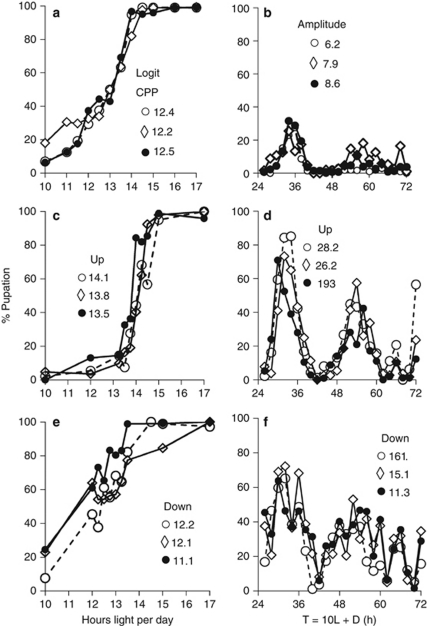
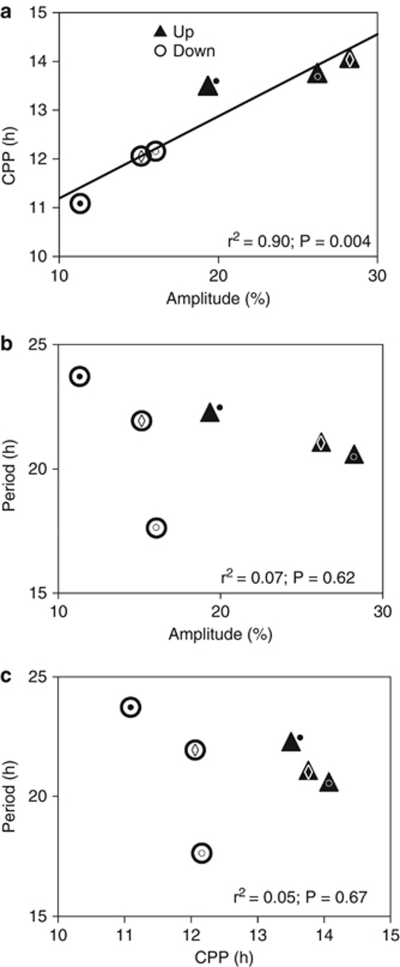
The genetic relationship between the daily circadian clock and the seasonal photoperiodic timer remains a subject of intense controversy. In Wyeomyia smithii, the critical photoperiod (an overt expression of the photoperiodic timer) evolves independently of the rhythmic response to the Nanda-Hamner protocol (an overt expression of the daily circadian clock) over a wide geographical range in North America. Herein, we focus on these two processes within a single local population in which there is a negative genetic correlation between them. We show that antagonistic selection against this genetic correlation rapidly breaks it down and, in fact, reverses its sign, showing that the genetic correlation is due primarily to linkage and not to pleiotropy. This rapid reversal of the genetic correlation within a small, single population means that it is difficult to argue that circadian rhythmicity forms the necessary, causal basis for the adaptive divergence of photoperiodic time measurement within populations or for the evolution of photoperiodic time measurement among populations over a broad geographical gradient of seasonal selection.
Figures
Comment in
-
What kind of insights can quantitative genetics provide us about this controversial hypothesis?Heredity (Edinb). 2012 May;108(5):469-70. doi: 10.1038/hdy.2011.121. Epub 2012 Feb 15. Heredity (Edinb). 2012. PMID: 22334116 Free PMC article. No abstract available.
References
-
- Bradshaw WE, Holzapfel CM. Evolution of animal photoperiodism. Annu Rev Ecol Evol Syst. 2007a;38:1–25.
-
- Bradshaw WE, Holzapfel CM. Tantalizing timeless. Science. 2007b;316:1851–1852. - PubMed
-
- Bradshaw WE, Holzapfel CM. What season is it anyway? Circadian tracking vs photoperiodic anticipation in insects. J Biol Rhythms. 2010a;25:155–165. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
