

Identification and characterization of Hoxa9 binding sites in hematopoietic cells
- PMID: 22072553
- PMCID: PMC3257007
- DOI: 10.1182/blood-2011-03-341081
Identification and characterization of Hoxa9 binding sites in hematopoietic cells
Abstract
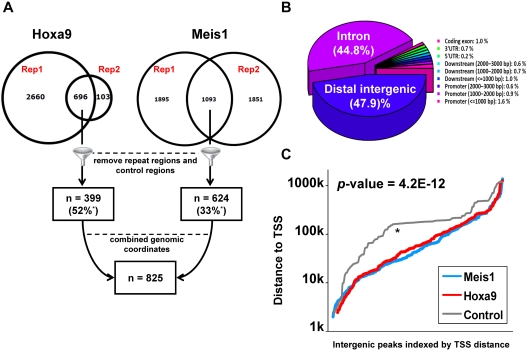
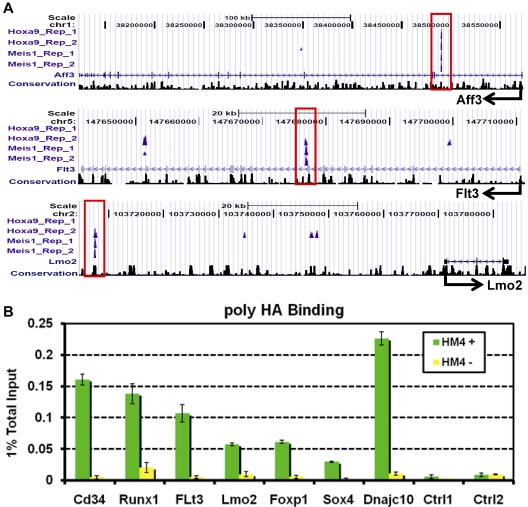
The clustered homeobox proteins play crucial roles in development, hematopoiesis, and leukemia, yet the targets they regulate and their mechanisms of action are poorly understood. Here, we identified the binding sites for Hoxa9 and the Hox cofactor Meis1 on a genome-wide level and profiled their associated epigenetic modifications and transcriptional targets. Hoxa9 and the Hox cofactor Meis1 cobind at hundreds of highly evolutionarily conserved sites, most of which are distant from transcription start sites. These sites show high levels of histone H3K4 monomethylation and CBP/P300 binding characteristic of enhancers. Furthermore, a subset of these sites shows enhancer activity in transient transfection assays. Many Hoxa9 and Meis1 binding sites are also bound by PU.1 and other lineage-restricted transcription factors previously implicated in establishment of myeloid enhancers. Conditional Hoxa9 activation is associated with CBP/P300 recruitment, histone acetylation, and transcriptional activation of a network of proto-oncogenes, including Erg, Flt3, Lmo2, Myb, and Sox4. Collectively, this work suggests that Hoxa9 regulates transcription by interacting with enhancers of genes important for hematopoiesis and leukemia.
Figures





References
-
- Argiropoulos B, Humphries RK. Hox genes in hematopoiesis and leukemogenesis. Oncogene. 2007;26(47):6766–6776. - PubMed
-
- Pineault N, Helgason CD, Lawrence HJ, Humphries RK. Differential expression of Hox, Meis1, and Pbx1 genes in primitive cells throughout murine hematopoietic ontogeny. Exp Hematol. 2002;30(1):49–57. - PubMed
-
- Thorsteinsdottir U, Mamo A, Kroon E, et al. Overexpression of the myeloid leukemia-associated Hoxa9 gene in bone marrow cells induces stem cell expansion. Blood. 2002;99(1):121–129. - PubMed
-
- Izon DJ, Rozenfeld S, Fong ST, Komuves L, Largman C, Lawrence HJ. Loss of function of the homeobox gene Hoxa-9 perturbs early T-cell development and induces apoptosis in primitive thymocytes. Blood. 1998;92(2):383–393. - PubMed
-
- Armstrong SA, Staunton JE, Silverman LB, et al. MLL translocations specify a distinct gene expression profile that distinguishes a unique leukemia. Nat Genet. 2002;30(1):41–47. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
