

Optimal control of natural eye-head movements minimizes the impact of noise
- PMID: 22072670
- PMCID: PMC6633248
- DOI: 10.1523/JNEUROSCI.3721-11.2011
Optimal control of natural eye-head movements minimizes the impact of noise
Abstract
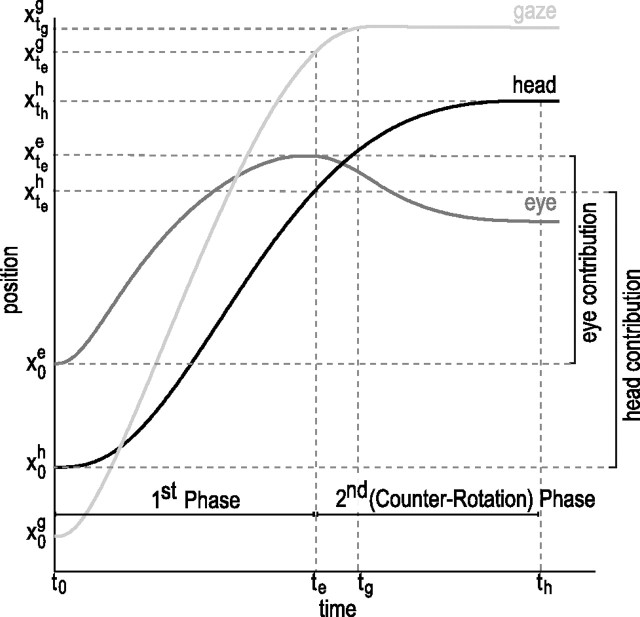
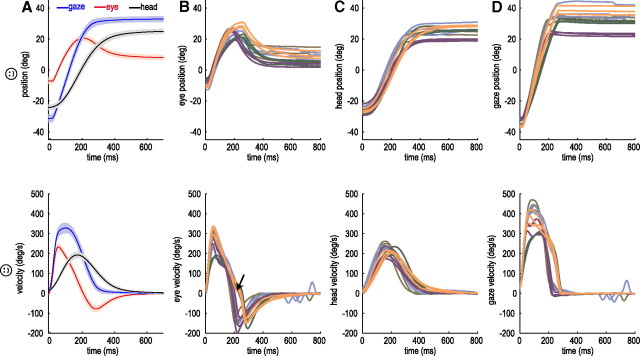
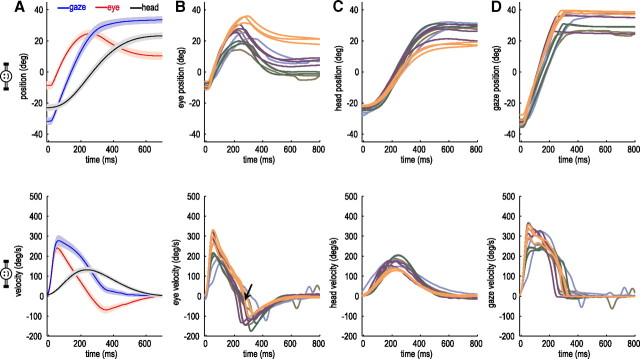
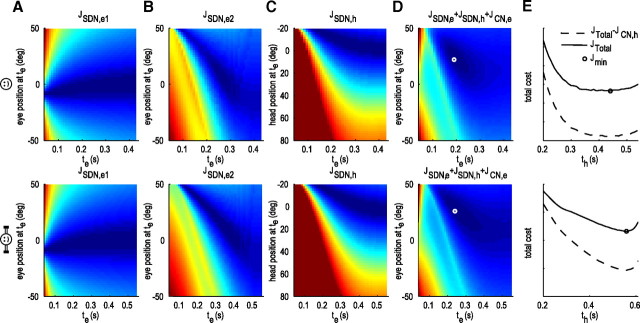
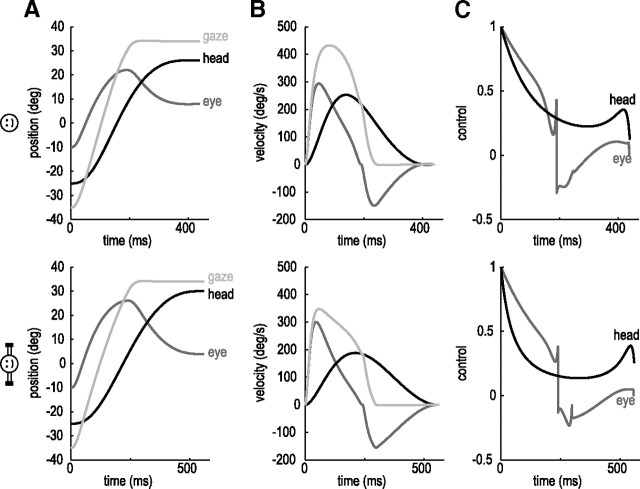
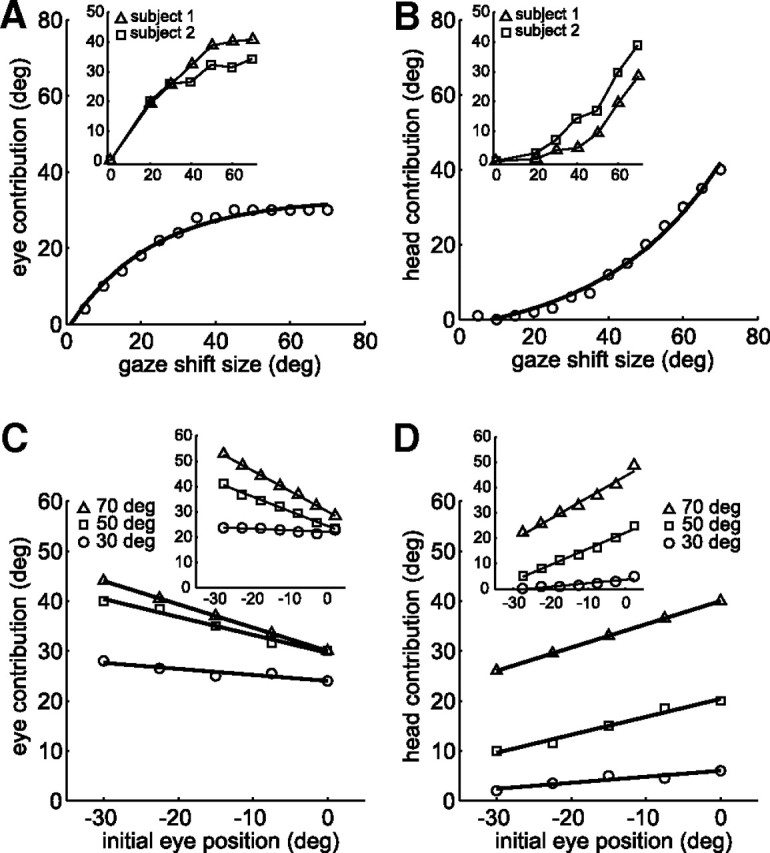
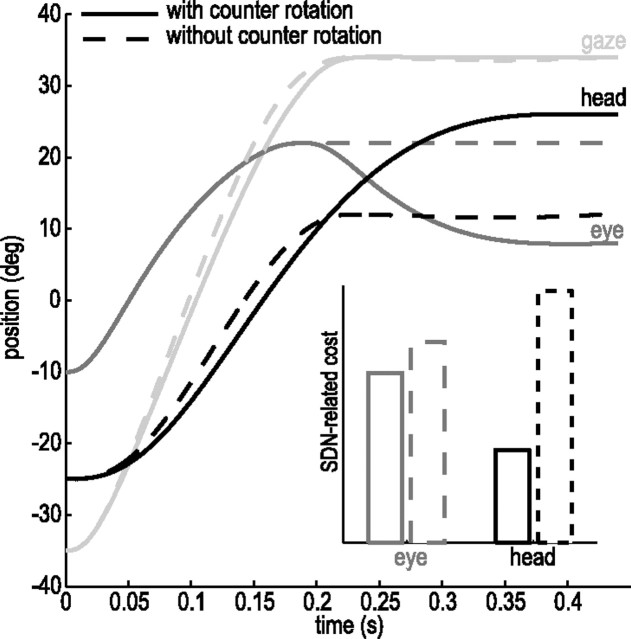
When shifting gaze to foveate a new target, humans mostly choose a unique set of eye and head movements from an infinite number of possible combinations. This stereotypy suggests that a general principle governs the movement choice. Here, we show that minimizing the impact of uncertainty, i.e., noise affecting motor performance, can account for the choice of combined eye-head movements. This optimization criterion predicts all major features of natural eye-head movements-including the part where gaze is already on target and the eye counter-rotates-such as movement durations, relative eye-head contributions, velocity profiles, and the dependency of gaze shifts on initial eye position. As a critical test of this principle, we show that it also correctly predicts changes in eye and head movement imposed by an experimental increase in the head moment of inertia. This suggests that minimizing the impact of noise is a simple and powerful principle that explains the choice of a unique set of movement profiles and segment coordination in goal-directed action.
Figures
References
-
- Freedman EG, Sparks DL. Eye-head coordination during head-unrestrained gaze shifts in rhesus monkeys. J Neurophysiol. 1997;77:2328–2348. - PubMed
-
- Freedman EG, Sparks DL. Coordination of the eyes and head: movement kinematics. Exp Brain Res. 2000;131:22–32. - PubMed
-
- Glasauer S. Cerebellar contribution to saccades and gaze holding: a modeling approach. Ann N Y Acad Sci. 2003;1004:206–219. - PubMed
-
- Glasauer S, Hoshi M, Kempermann U, Eggert T, Büttner U. Three-dimensional eye position and slow phase velocity in humans with downbeat nystagmus. J Neurophysiol. 2003;89:338–354. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical