

A novel protein LZTFL1 regulates ciliary trafficking of the BBSome and Smoothened
- PMID: 22072986
- PMCID: PMC3207910
- DOI: 10.1371/journal.pgen.1002358
A novel protein LZTFL1 regulates ciliary trafficking of the BBSome and Smoothened
Abstract
Many signaling proteins including G protein-coupled receptors localize to primary cilia, regulating cellular processes including differentiation, proliferation, organogenesis, and tumorigenesis. Bardet-Biedl Syndrome (BBS) proteins are involved in maintaining ciliary function by mediating protein trafficking to the cilia. However, the mechanisms governing ciliary trafficking by BBS proteins are not well understood. Here, we show that a novel protein, Leucine-zipper transcription factor-like 1 (LZTFL1), interacts with a BBS protein complex known as the BBSome and regulates ciliary trafficking of this complex. We also show that all BBSome subunits and BBS3 (also known as ARL6) are required for BBSome ciliary entry and that reduction of LZTFL1 restores BBSome trafficking to cilia in BBS3 and BBS5 depleted cells. Finally, we found that BBS proteins and LZTFL1 regulate ciliary trafficking of hedgehog signal transducer, Smoothened. Our findings suggest that LZTFL1 is an important regulator of BBSome ciliary trafficking and hedgehog signaling.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
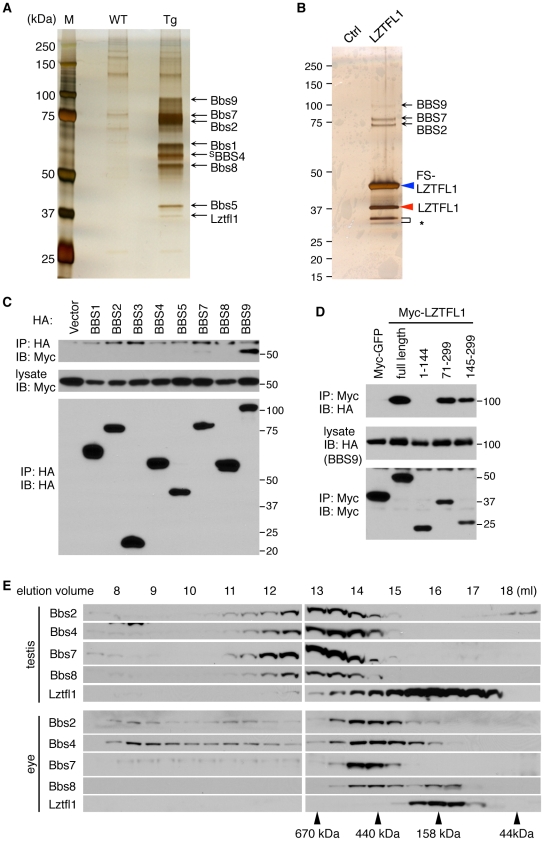
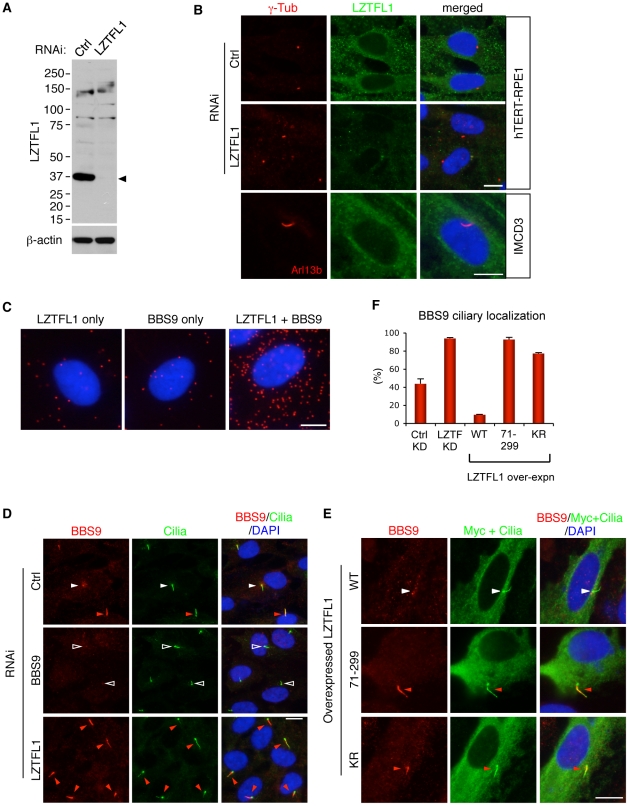
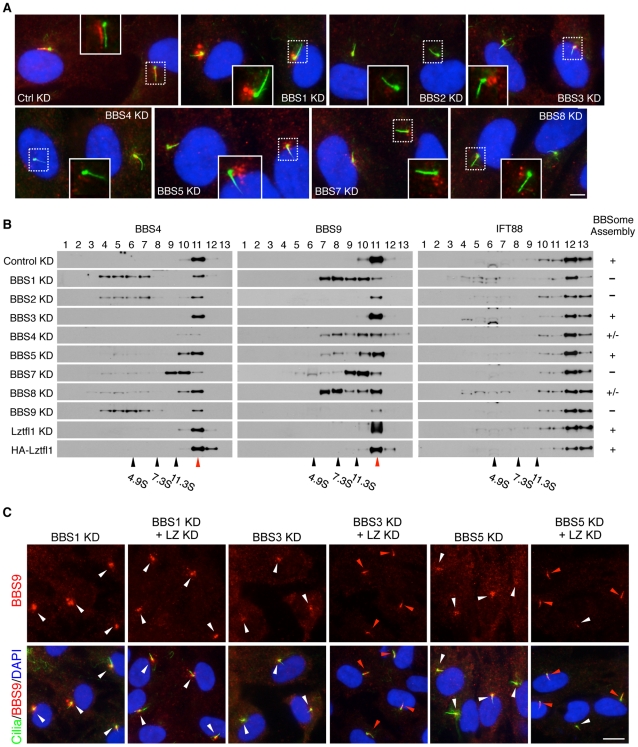
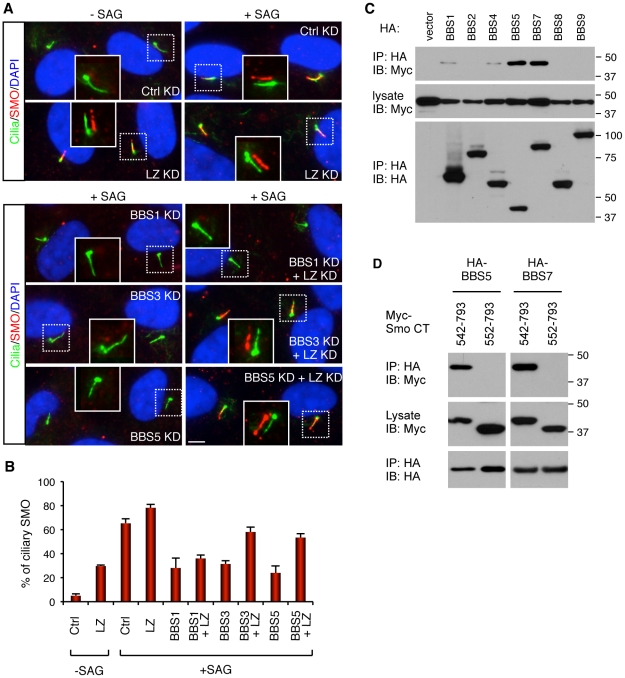
References
-
- Singla V, Reiter JF. The primary cilium as the cell's antenna: signaling at a sensory organelle. Science. 2006;313:629–633. - PubMed
-
- Quinlan RJ, Tobin JL, Beales PL. Modeling ciliopathies: Primary cilia in development and disease. Curr Top Dev Biol. 2008;84:249–310. - PubMed
-
- Badano JL, Mitsuma N, Beales PL, Katsanis N. The ciliopathies: an emerging class of human genetic disorders. Annu Rev Genomics Hum Genet. 2006;7:125–148. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
