

Review
doi: 10.1074/jbc.R111.303990.
Epub 2011 Nov 10.
Metabolism of carotenoids and retinoids related to vision
Affiliations
- PMID: 22074927
- PMCID: PMC3265843
- DOI: 10.1074/jbc.R111.303990
Item in Clipboard
Review
Metabolism of carotenoids and retinoids related to vision
J Biol Chem.
.
Abstract
All animals endowed with the ability to detect light through visual pigments must have evolved pathways in which dietary precursors for the involved chromophore are absorbed, transported, and metabolized. Knowledge about this metabolism has exponentially increased over the past decade. Genetic manipulation of animal models provided insights into the metabolic flow of these compounds through the body and in the eyes, unraveling their regulatory aspects and aberrant side reactions. The scheme that emerges reveals a common origin of key components for chromophore metabolism that have been adapted to the specific requirements of retinoid biology in different animal classes.
Figures
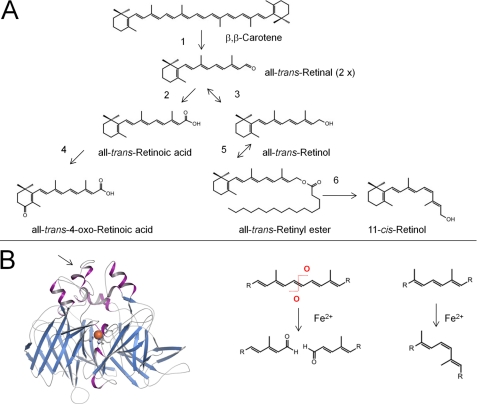
Carotenoids and retinoids undergo limited number of chemical reactions. A, enzymatic steps in vertebrate carotenoid/retinoid metabolism. Note that similar enzymatic modifications of carotenoids and apocarotenoids occur in plants, fungi, and bacteria, and these are catalyzed by related enzymes. Step 1, oxidative cleavage of double bonds; steps 2 and 3, oxidation and reduction of oxygen end groups; step 4, introduction of oxygen into the ionone ring; step 5, esterification of hydroxyl groups; step 6, trans-to-cis isomerization of double bonds of the polyene chain. B, left, the crystal structure of RPE65 (retinoid isomerase) from Bos taurus is shown. The structural fold is well conserved in plant and bacterial CCEs. The arrow points to a region of RPE65 that associates with the lipid membrane. The ferrous iron that demarks the reaction center of this type of protein is highlighted as an orange sphere. Right, the enzymatic reactions catalyzed by this class of enzymes include oxidative cleavage in and trans-to-cis isomerization of double bonds of the polyene chain of carotenoids and apocarotenoids.

Carotenoid uptake is protein-facilitated process. A, in animals, class B scavenger receptors such as NinaB, SANTA MARIA, and SR-BI facilitate the cellular uptake of carotenoids from micelles in the gut and/or circulating lipoproteins in the blood. Structural prediction based on the primary amino acid sequence of this class of proteins indicates that these receptors possess two membrane-spanning domains at the N and C termini as well as a large extracellular loop. B, upon cellular absorption, carotenoids can be sequestered by binding proteins and lipid droplets. Additionally, carotenoids can be converted to apocarotenoids, including retinoids, by carotenoid oxygenases. Genetic disruption of these processes alters carotenoid homeostasis. Left panel, mutations in the Yellow Cocoon gene encoding a class B scavenger receptor or mutations in the Yellow Blood gene encoding a carotenoid-binding protein alter silk color in B. mori. Genetic disruption of Bcmo1 results in β,β-carotene accumulation in the fat pads of mice. Mutations in the ninaB gene lead to carotenoid accumulation in the eye. Genetic disruption of the Bcdo2 gene results in a yellow color of isolated liver mitochondria due to carotenoid accumulation.
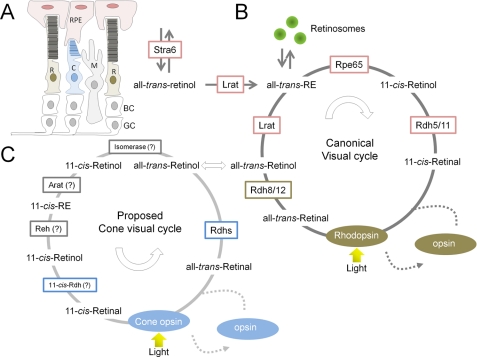
Retinoid metabolism in mammalian eyes. A, simplified schematic overview of the mammalian retina. R, rod photoreceptors; C, cone photoreceptors; M, Müller glial cells. Second-order bipolar cells (BC) and third-order ganglion cells (GC) are shown but not discussed. The drawing is adapted from Ref. . B, biochemical key steps of the canonical visual cycle. C, proposed biochemical steps of the alternative visual cycle for rods. Different enzymes involved in the canonical and cone-specific visual cycles are indicated. Enzymes that have not been molecularly identified are denoted by question marks. The colored boxes enclosing the enzyme names indicate their cellular localization: pink, RPE; green, ROS; blue, cone outer segments; gray, Müller glial cells. Note that transport of retinoids in and between cells requires binding proteins such as intracellular and extracellular RBPs as well as the putative retinoid transporter ABCA4. Reh, RE hydrolase; Arat, acetyl-CoA:retinol acyltransferase.
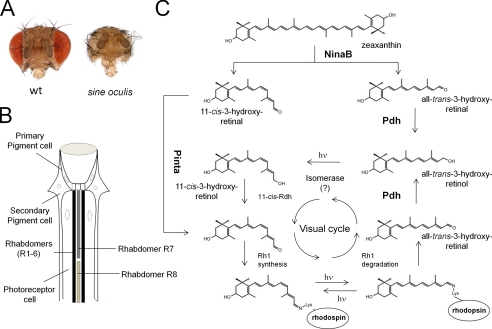
Ocular carotenoid/retinoid metabolism in Drosophila. A, images of the heads of wild-type flies and sine oculis flies, which lack compound eyes. B, schematic drawing of a longitudinal section through a fly omatidium. The visual cycle of the fly operates between the photoreceptor cells and the secondary pigment cells. Visual pigments are sequestered in rhabdomeres R1–R6 of photoreceptor cells, which express ninaE, and rhabdomers R7 and R8, which express other opsin genes. C, zeaxanthin is converted to one molecule of 11-cis-3-hydroxyretinal and one molecule of all-trans-3-hydroxyretinal by NinaB expressed in neuronal and glial cells. Transport of 11-cis-3-hydroxyretinal to photoreceptors depends on the PINTA protein. The all-trans-3-hydroxyretinal cleavage product is converted to all-trans-3-hydroxyretinol by PDH. A blue light-dependent isomerase reaction mediates 11-cis-3-hydroxyretinol production. 11-cis-3-Hydroxyretinol is then converted to 11-cis-3-hydroxyretinal. The 11-cis-chromophore binds to different fly opsins to form functional visual pigments. Insect visual pigments are bistable, so the opsin-bound chromophore can be isomerized back to the 11-cis geometric state by light. However, a regeneration pathway for the chromophore is required for the released chromophore from internalized rhodopsin. This all-trans-3-hydroxyretinal is converted by PDH into all-trans-3-hydroxyretinol, which can be recycled back to the chromophore by the enzymatic steps described above. hν, one photon.
Similar articles
-
Molecular components affecting ocular carotenoid and retinoid homeostasis.Prog Retin Eye Res. 2021 Jan;80:100864. doi: 10.1016/j.preteyeres.2020.100864. Epub 2020 Apr 25. Prog Retin Eye Res. 2021. PMID: 32339666 Free PMC article. Review.
-
The molecular aspects of absorption and metabolism of carotenoids and retinoids in vertebrates.Biochim Biophys Acta Mol Cell Biol Lipids. 2020 Nov;1865(11):158571. doi: 10.1016/j.bbalip.2019.158571. Epub 2019 Nov 23. Biochim Biophys Acta Mol Cell Biol Lipids. 2020. PMID: 31770587 Free PMC article. Review.
-
Confronting complexity: the interlink of phototransduction and retinoid metabolism in the vertebrate retina.Prog Retin Eye Res. 2001 Jul;20(4):469-529. doi: 10.1016/s1350-9462(01)00002-7. Prog Retin Eye Res. 2001. PMID: 11390257 Review.
-
Carotenoid modifying enzymes in metazoans.Methods Enzymol. 2022;674:405-445. doi: 10.1016/bs.mie.2022.05.005. Epub 2022 Jun 22. Methods Enzymol. 2022. PMID: 36008015 Free PMC article.
-
Evolution of the visual cycle: the role of retinoid-binding proteins.J Endocrinol. 2002 Oct;175(1):75-88. doi: 10.1677/joe.0.1750075. J Endocrinol. 2002. PMID: 12379492 Review.
Cited by
-
Mapping of the extracellular RBP4 ligand binding domain on the RBPR2 receptor for Vitamin A transport.Front Cell Dev Biol. 2023 Feb 22;11:1105657. doi: 10.3389/fcell.2023.1105657. eCollection 2023. Front Cell Dev Biol. 2023. PMID: 36910150 Free PMC article.
-
Molecular aspects of β, β-carotene-9', 10'-oxygenase 2 in carotenoid metabolism and diseases.Exp Biol Med (Maywood). 2016 Nov;241(17):1879-1887. doi: 10.1177/1535370216657900. Epub 2016 Jul 7. Exp Biol Med (Maywood). 2016. PMID: 27390265 Free PMC article. Review.
-
Consumption of a high-fat diet alters transcriptional rhythmicity in liver from pubertal mice.Front Nutr. 2023 Jan 4;9:1068350. doi: 10.3389/fnut.2022.1068350. eCollection 2022. Front Nutr. 2023. PMID: 36687679 Free PMC article.
-
Evidence for compartmentalization of mammalian carotenoid metabolism.FASEB J. 2014 Oct;28(10):4457-69. doi: 10.1096/fj.14-252411. Epub 2014 Jul 7. FASEB J. 2014. PMID: 25002123 Free PMC article.
-
Transport of vitamin A across blood-tissue barriers is facilitated by STRA6.FASEB J. 2016 Aug;30(8):2985-95. doi: 10.1096/fj.201600446R. Epub 2016 May 17. FASEB J. 2016. PMID: 27189978 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
