

doi: 10.1016/j.cub.2011.09.040.
Copper: an essential metal in biology
Affiliations
- PMID: 22075424
- PMCID: PMC3718004
- DOI: 10.1016/j.cub.2011.09.040
Item in Clipboard
Copper: an essential metal in biology
Curr Biol.
.
No abstract available
Figures

The themes of Cu homeostasis are conserved among Kingdoms. Import (orange), chaperones (blue), export (yellow), transcriptional control (grey). The general bacterial Cu response activates an operon consisting of a transcriptional repressor, a chaperone and a P-type exporter. In the absence of Cu, transcriptional repressors such as CueR, CsoR and CopY maintain repression of the Cu response machinery. In the presence of Cu, repressor proteins bind Cu and release from their promoter DNA bindings sites, resulting in derepression of genes encoding chaperones and P-type ATPases to facilitate protection against Cu toxicity by sequestration and export. Cytochrome c oxidase, located in the bacterial membrane, is the most common cuproenzyme in bacteria. The mechanism of Cu import in bacterial cells is currently poorly defined but may include a relatively non-metal-specific cation transporter. Alternatively, methanotrophic bacteria may use a non-translationally synthesized Cu binding peptide, methanobactin, to acquire Cu.
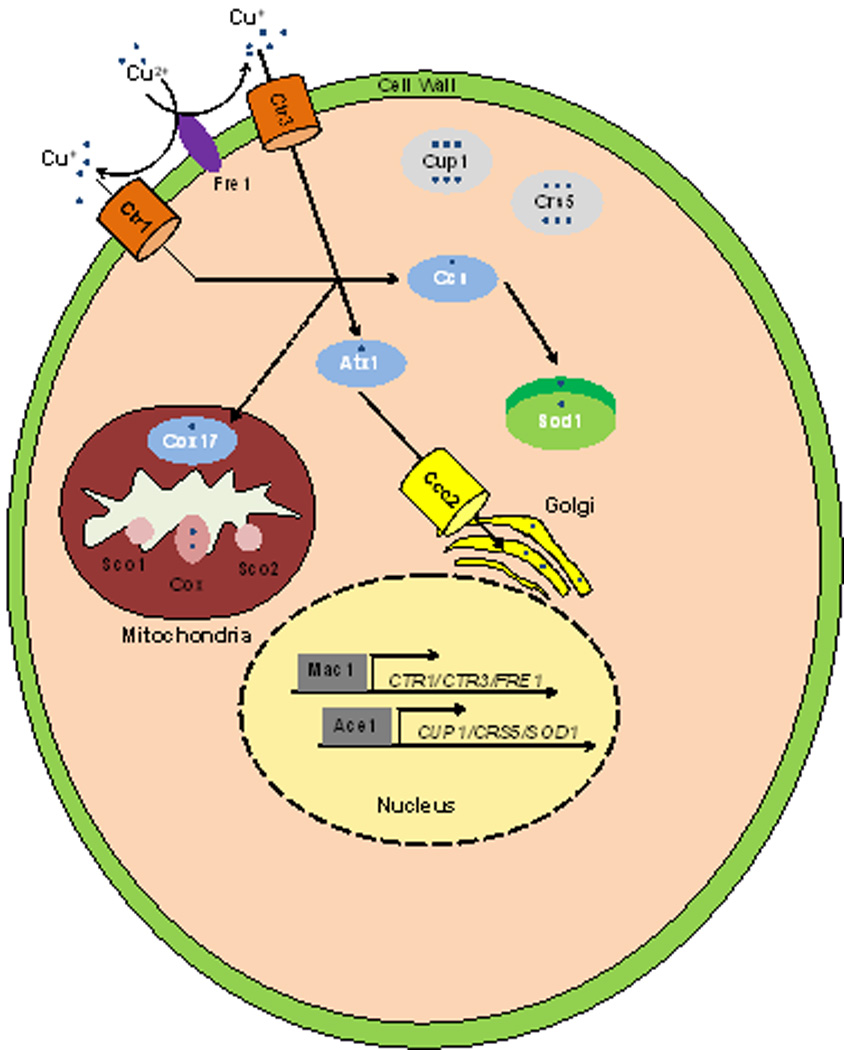
Unicellular organisms such as yeast have increased complexity to distribute Cu to intracellular proteins and compartments. Most notably, chaperones are utilized to safeguard the cell from free Cu and to expedite delivery, as the metal is delivered to intracellular Cu containing proteins. Ccs delivers Cu after reduction from Cu2+ to Cu+ by the Fre1 metalloreductase, and imported by the Ctr1 and Ctr3 Cu transporters, to Sod1 to protect against oxidative stress. Atx1 is a chaperone that delivers Cu to Ccc2 where Cu is translocated into the Golgi apparatus to be loaded onto cuproenzyems. Cup1 and Crs5 are metallothioneins that protect yeast cells from Cu toxicity. Mac1 and Ace1 are transcriptional activators that respond to Cu starvation and excess respectively. The precise mechanisms whereby cytosolic Cu is delivered to the mitochondria, for loading onto cytochrome oxidase (Cox) by Cox17, Sco1/2 and other proteins, is currently unknown.
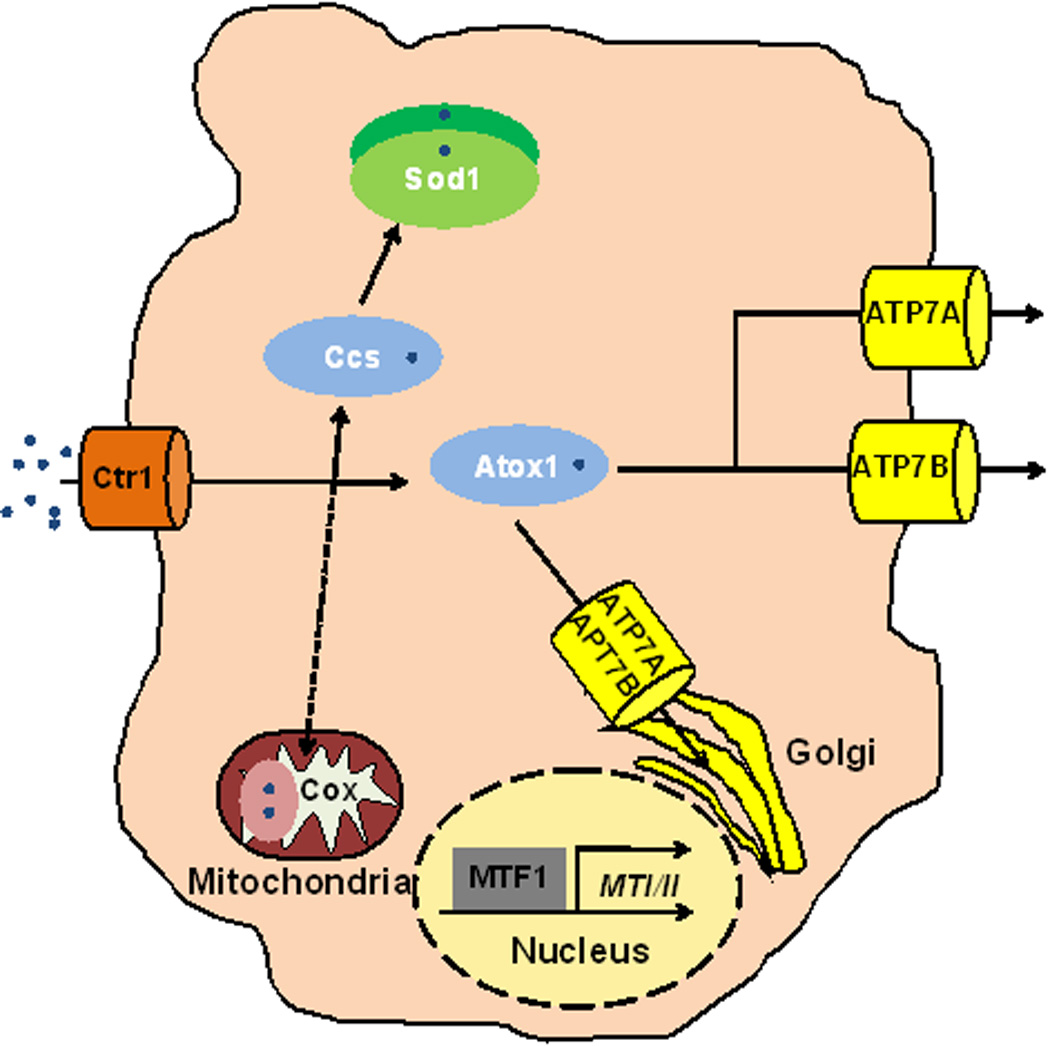
A generalized mammalian cell retains similar Cu homeostasis machinery to yeast, with increased complexity of components. Ctr1 is the predominant importer of Cu into mammalian cells. The Ccs and Sod1 chaperones are conserved from single cell eukaryotes. Atox1, the Atx1 homologue in mammals is responsible for delivery of Cu to the two P-type ATPases in mammals, ATP7A and ATP7B. These two P-type ATPases are also important for delivery of Cu to the Golgi to load onto proteins and in distinct cell types such as intestinal epithelial cells or hepatocytes, to efflux Cu. MTF1 transcriptionally activates metallothionein genes (MTI/II) and other targets. How Cu status is communicated between tissues must be explored to understand Cu homeostasis throughout metazoan organisms.
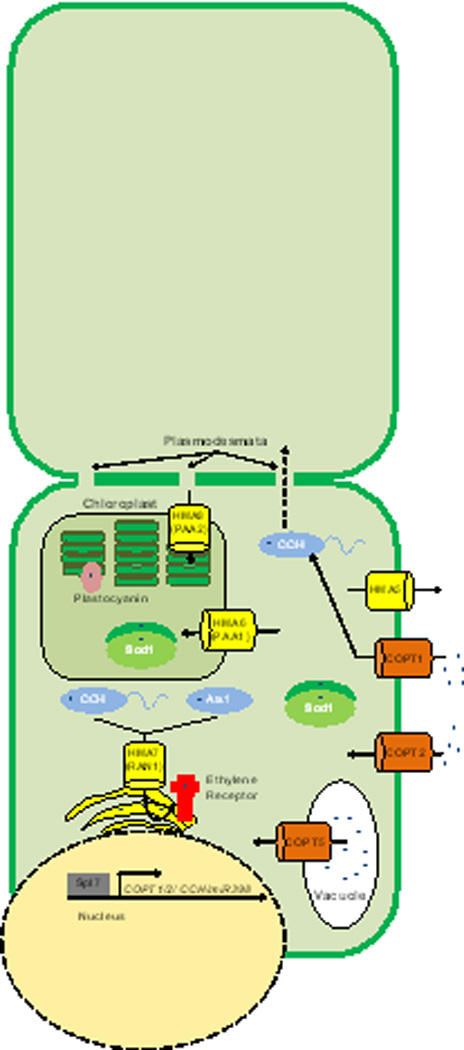
Plants have expanded the number of encoded Cu homeostasis proteins. The COPT proteins are similar to the Ctr transporters found in mammals and yeasts. COPT1 imports Cu and delivers it to CCH. CCH, while similar to Atox1, has a carboxy-terminal extension that may be important for cell-to-cell transport of Cu between plasmodesmata. Plants do encode another Atox1 homologue, ATX1. Copt5 has been associated with Cu export from the vacuole during times of Cu starvation. Chloroplast specific importers (PAA1 and PAA2) have also been identified and help to deliver Cu to important chloroplast specific proteins such as plastocyanin. RAN1 plays a similar role to Ccc2 by transporting Cu to the Golgi to be loaded onto nascent proteins such as the ethylene receptor. Spl7 is a Cu-responsive transcriptional regulator that is responsible for activation and expression of CCH, COPT1, and COPT2 among other genes, as well as micro RNAs that impact the expression of other Cu related genes such as that encoding Cu, Zn SOD1.
References
-
- Kim BE, Nevitt T, Thiele DJ. Mechanisms for copper acquisition, distribution and regulation. Nat Chem Biol. 2008;4:176–185. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
