

Alkamides activate jasmonic acid biosynthesis and signaling pathways and confer resistance to Botrytis cinerea in Arabidopsis thaliana
- PMID: 22076141
- PMCID: PMC3208606
- DOI: 10.1371/journal.pone.0027251
Alkamides activate jasmonic acid biosynthesis and signaling pathways and confer resistance to Botrytis cinerea in Arabidopsis thaliana
Abstract
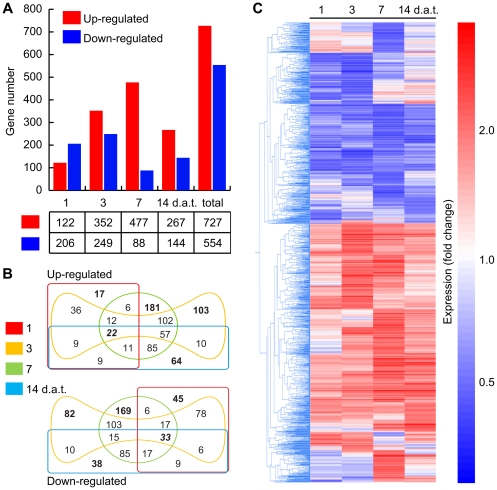
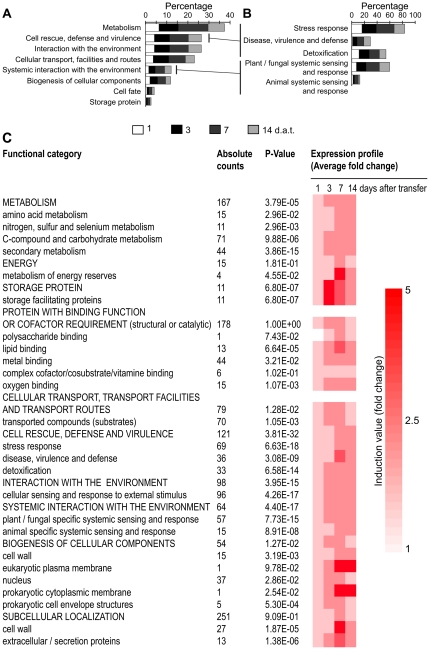
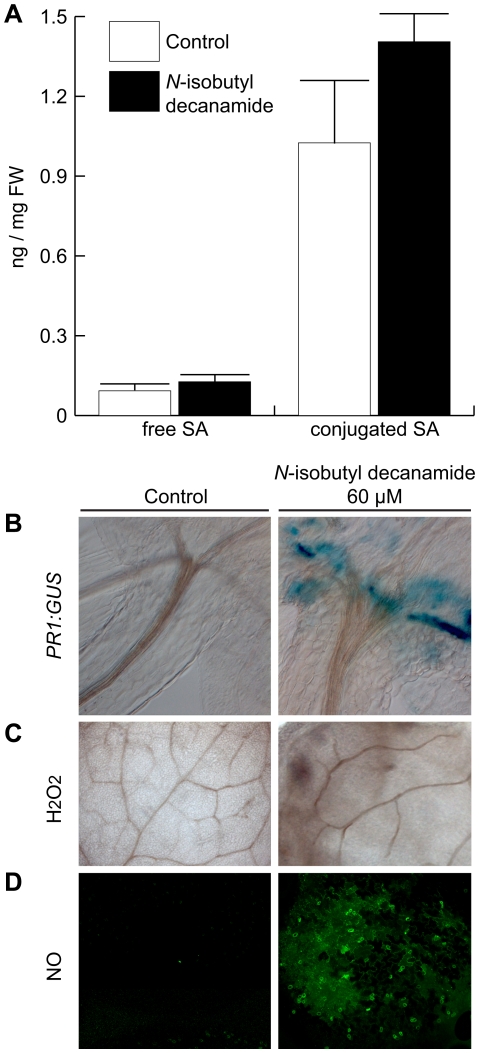
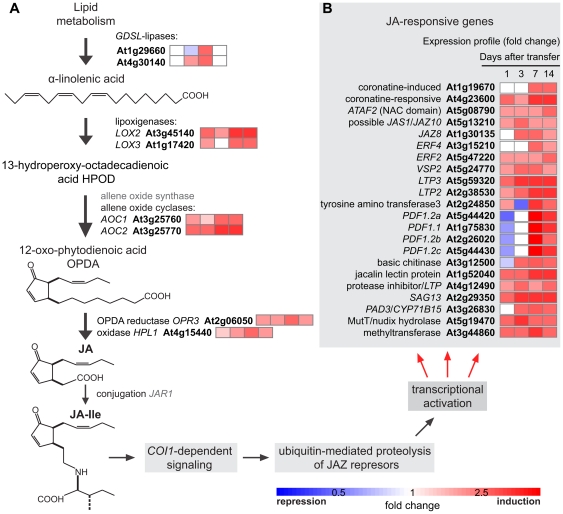
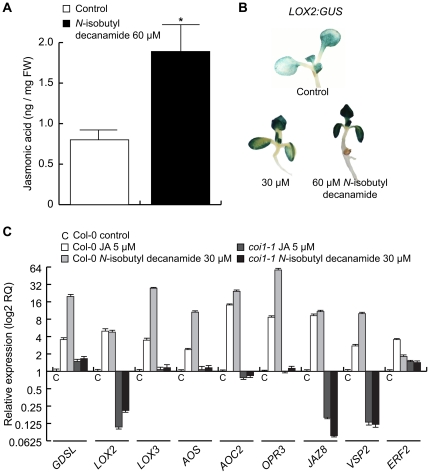
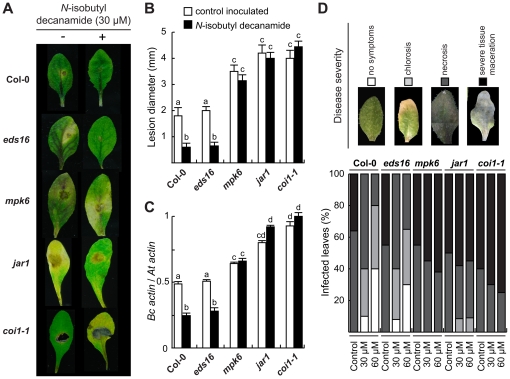
Alkamides are fatty acid amides of wide distribution in plants, structurally related to N-acyl-L-homoserine lactones (AHLs) from Gram-negative bacteria and to N- acylethanolamines (NAEs) from plants and mammals. Global analysis of gene expression changes in Arabidopsis thaliana in response to N-isobutyl decanamide, the most highly active alkamide identified to date, revealed an overrepresentation of defense-responsive transcriptional networks. In particular, genes encoding enzymes for jasmonic acid (JA) biosynthesis increased their expression, which occurred in parallel with JA, nitric oxide (NO) and H₂O₂ accumulation. The activity of the alkamide to confer resistance against the necrotizing fungus Botrytis cinerea was tested by inoculating Arabidopsis detached leaves with conidiospores and evaluating disease symptoms and fungal proliferation. N-isobutyl decanamide application significantly reduced necrosis caused by the pathogen and inhibited fungal proliferation. Arabidopsis mutants jar1 and coi1 altered in JA signaling and a MAP kinase mutant (mpk6), unlike salicylic acid- (SA) related mutant eds16/sid2-1, were unable to defend from fungal attack even when N-isobutyl decanamide was supplied, indicating that alkamides could modulate some necrotrophic-associated defense responses through JA-dependent and MPK6-regulated signaling pathways. Our results suggest a role of alkamides in plant immunity induction.
Conflict of interest statement
Figures

References
-
- Glazebrook J. Genes controlling expression of defense responses in Arabidopsis - 2001 status. Curr Opin Plant Biol. 2001;4:301–308. - PubMed
-
- Wendehenne D, Courtois C, Besson A, Gravot A, Buchwalter A, et al. Lamattina L, Polacco JC, editors. NO-based signaling in plants. Nitric oxide in plant growth. 2006. pp. 35–51. Plant Cell Monogr vol 6. Springer-Verlag, Berlin Heidelberg New York.
-
- Satner A, Estelle M. Recent advances and emerging trends in plant hormone signaling. Nature. 2009;459:1071–1078. - PubMed
-
- Shah J. Lipids, lipases, and lipid-modifying enzymes in plant disease resistance. Annu Rev Phytopathol. 2005;43:229–260. - PubMed
-
- Upchurch RG. Fatty acid unsaturation, mobilization, and regulation in the response of plants to stress. Biotech Lett. 2008;30(6):967–977. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
