

Predicting the potential demographic impact of predators on their prey: a comparative analysis of two carnivore-ungulate systems in Scandinavia
- PMID: 22077484
- PMCID: PMC3440569
- DOI: 10.1111/j.1365-2656.2011.01928.x
Predicting the potential demographic impact of predators on their prey: a comparative analysis of two carnivore-ungulate systems in Scandinavia
Abstract
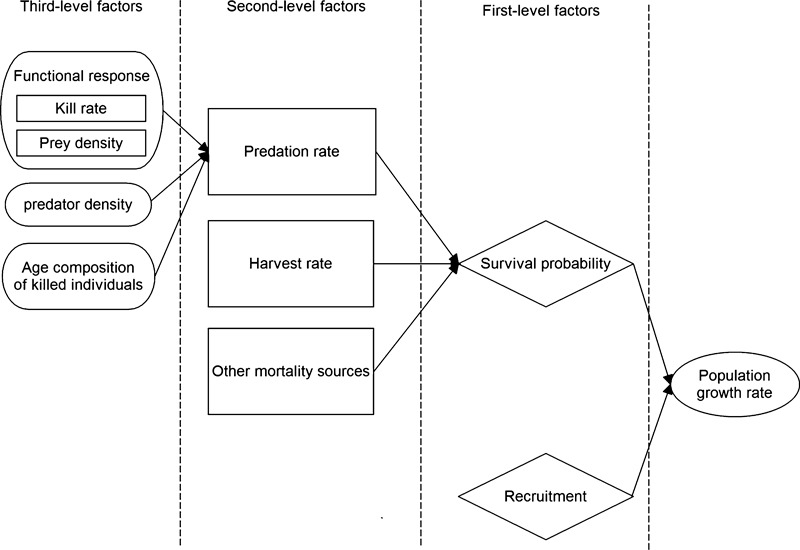
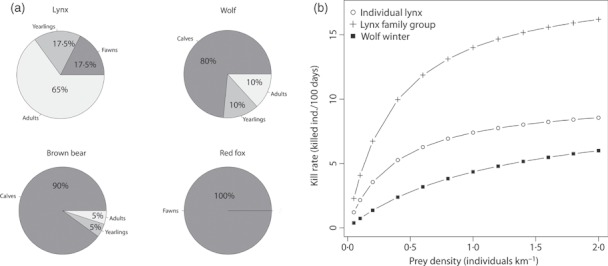
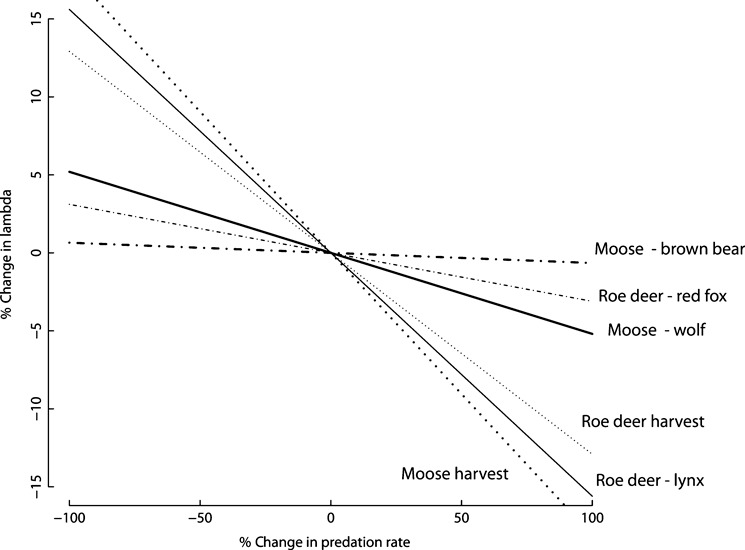
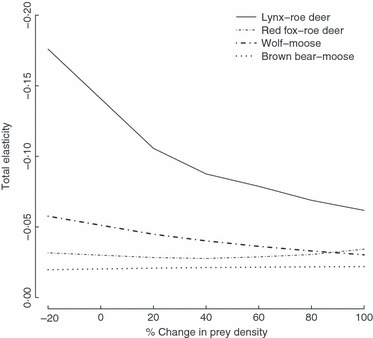
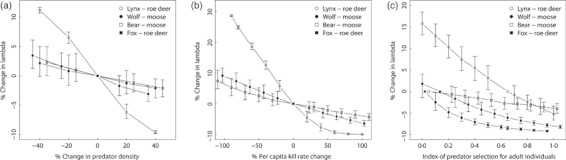
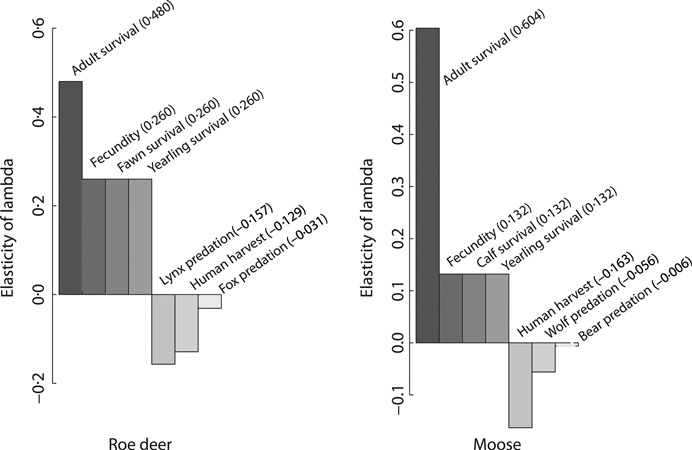
1. Understanding the role of predation in shaping the dynamics of animal communities is a fundamental issue in ecological research. Nevertheless, the complex nature of predator-prey interactions often prevents researchers from modelling them explicitly. 2. By using periodic Leslie-Usher matrices and a simulation approach together with parameters obtained from long-term field projects, we reconstructed the underlying mechanisms of predator-prey demographic interactions and compared the dynamics of the roe deer-red fox-Eurasian lynx-human harvest system with those of the moose-brown bear-gray wolf-human harvest system in the boreal forest ecosystem of the southern Scandinavian Peninsula. 3. The functional relationship of both roe deer and moose λ to changes in predation rates from the four predators was remarkably different. Lynx had the strongest impact among the four predators, whereas predation rates by wolves, red foxes, or brown bears generated minor variations in prey population λ. Elasticity values of lynx, wolf, fox and bear predation rates were -0·157, -0·056, -0·031 and -0·006, respectively, but varied with both predator and prey densities. 4. Differences in predation impact were only partially related to differences in kill or predation rates, but were rather a result of different distribution of predation events among prey age classes. Therefore, the age composition of killed individuals emerged as the main underlying factor determining the overall per capita impact of predation. 5. Our results confirm the complex nature of predator-prey interactions in large terrestrial mammals, by showing that different carnivores preying on the same prey species can exert a dramatically different demographic impact, even in the same ecological context, as a direct consequence of their predation patterns. Similar applications of this analytical framework in other geographical and ecological contexts are needed, but a more general evaluation of the subject is also required, aimed to assess, on a broader systematic and ecological range, what specific traits of a carnivore are most related to its potential impact on prey species.
© 2011 The Authors. Journal of Animal Ecology © 2011 British Ecological Society.
Figures
References
-
- Abrams P, Ginzburg L. The nature of predation: prey dependent, ratio dependent or neither? Trends in Ecology and Evolution. 2000;15:337–341. - PubMed
-
- Andersen R, Linnell JDC. Irruptive potential in roe deer: density dependent effects on rate of increase and fecundity. Journal of Wildlife Management. 2000;64:698–706.
-
- Andersen R, Karlsen J, Austmo LB, Odden J, Linnell JDC, Gaillard JM. Selectivity of Eurasian lynx Lynx lynx and recreational hunters for age, sex and body condition in roe deer Capreolus capreolus. Wildlife Biology. 2007;13:467–474.
-
- Atwood TC, Gese EM, Kunkel KE. Comparative patterns of predation by cougars and recolonizing wolves in Montana's Madison Range. Journal of Wildlife Management. 2007;71:1098–1106.
-
- Ballard WB, Whitman JS, Gardner CL. Ecology of an exploited wolf population in south-central Alaska. Wildlife Monographs. 1987;54:1–54.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
