

Systematic discovery of TLR signaling components delineates viral-sensing circuits
- PMID: 22078882
- PMCID: PMC3809888
- DOI: 10.1016/j.cell.2011.10.022
Systematic discovery of TLR signaling components delineates viral-sensing circuits
Abstract
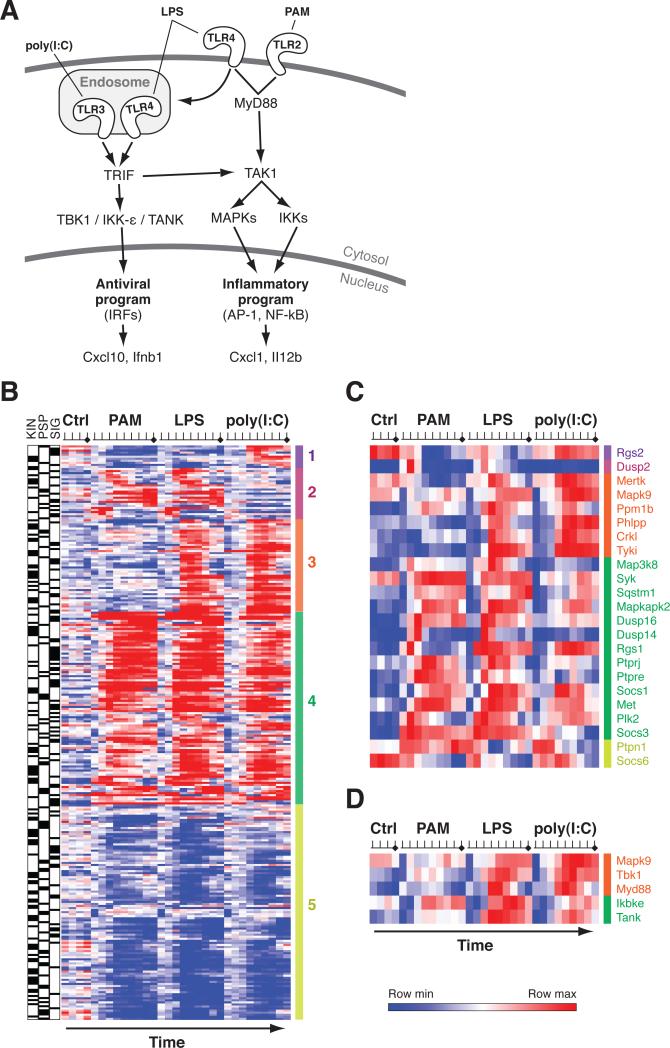
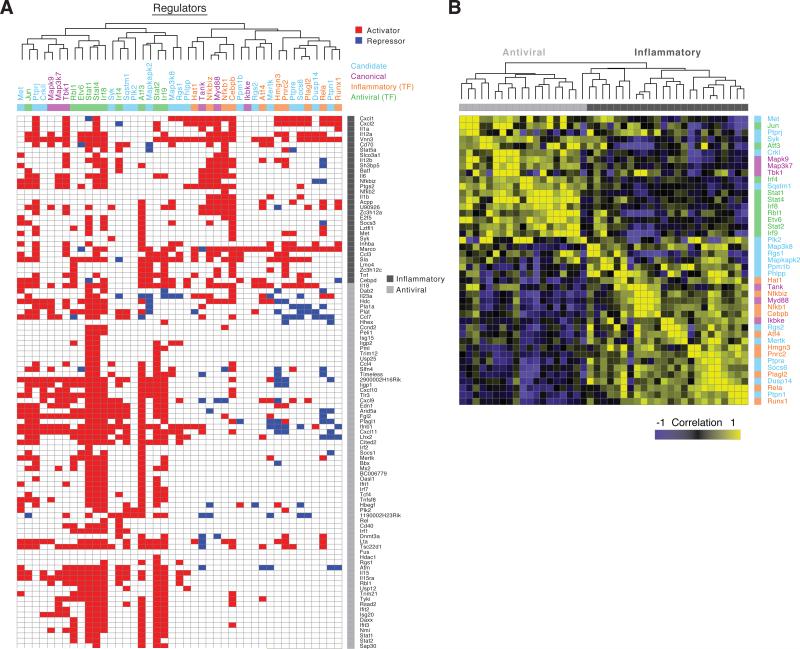
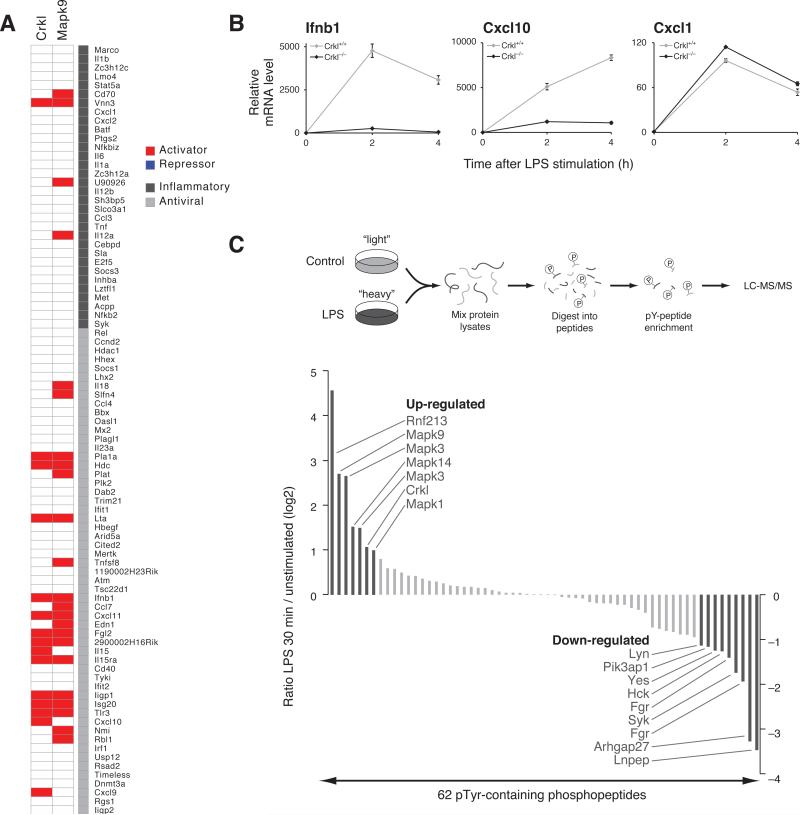
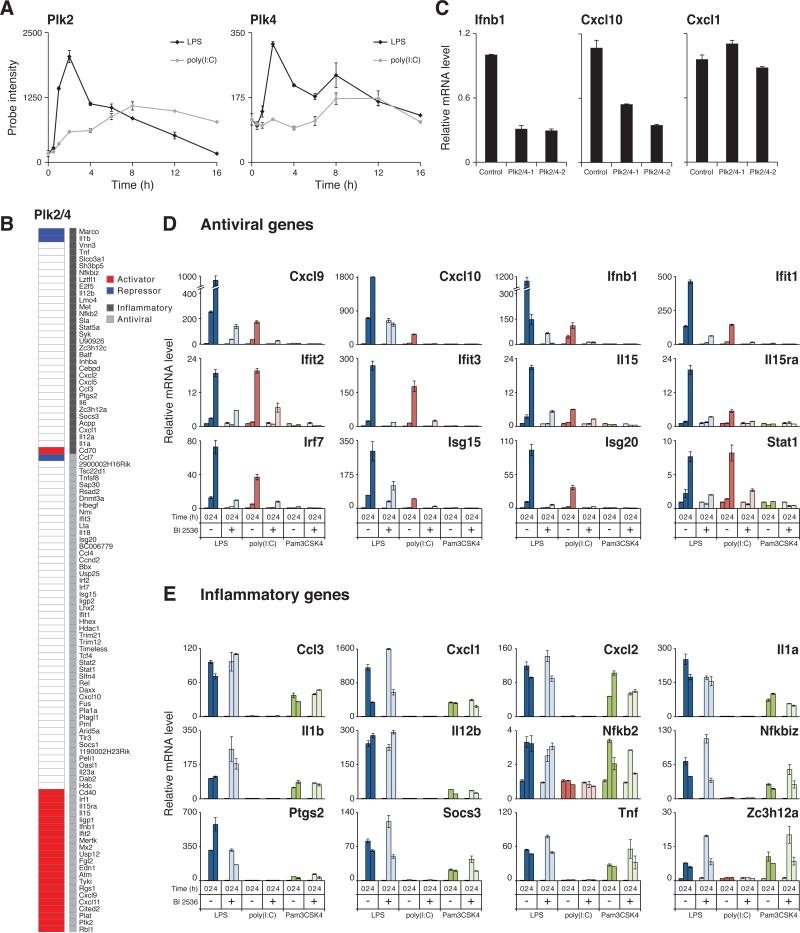
Deciphering the signaling networks that underlie normal and disease processes remains a major challenge. Here, we report the discovery of signaling components involved in the Toll-like receptor (TLR) response of immune dendritic cells (DCs), including a previously unkown pathway shared across mammalian antiviral responses. By combining transcriptional profiling, genetic and small-molecule perturbations, and phosphoproteomics, we uncover 35 signaling regulators, including 16 known regulators, involved in TLR signaling. In particular, we find that Polo-like kinases (Plk) 2 and 4 are essential components of antiviral pathways in vitro and in vivo and activate a signaling branch involving a dozen proteins, among which is Tnfaip2, a gene associated with autoimmune diseases but whose role was unknown. Our study illustrates the power of combining systematic measurements and perturbations to elucidate complex signaling circuits and discover potential therapeutic targets.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
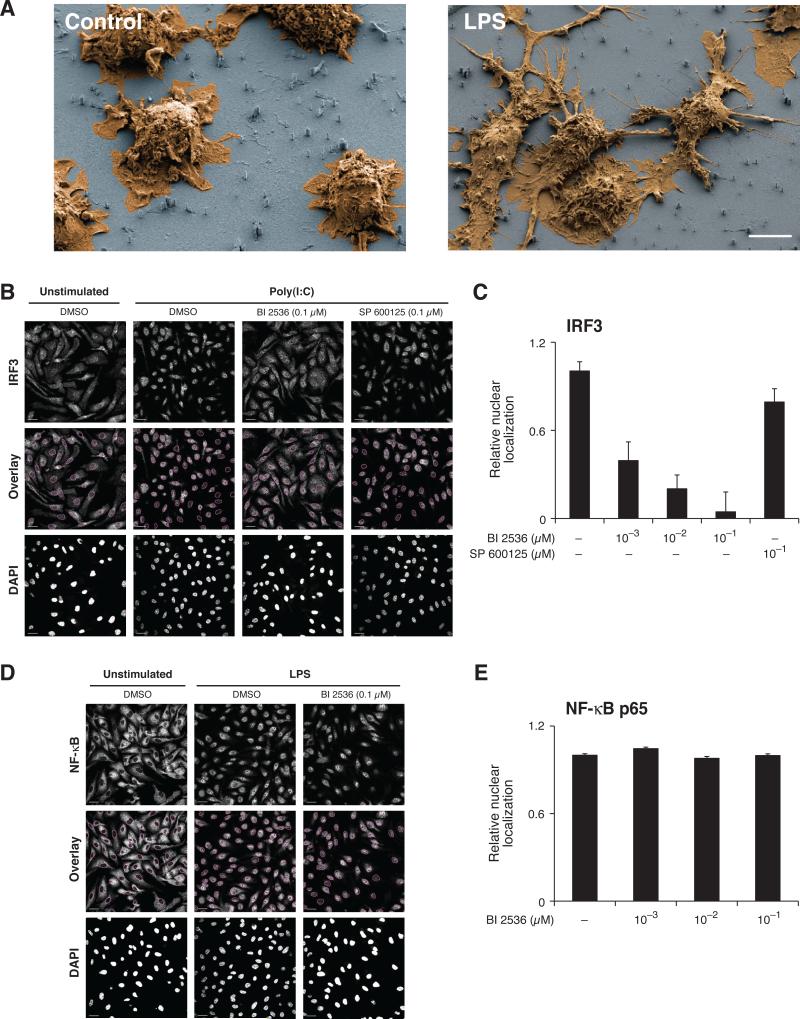
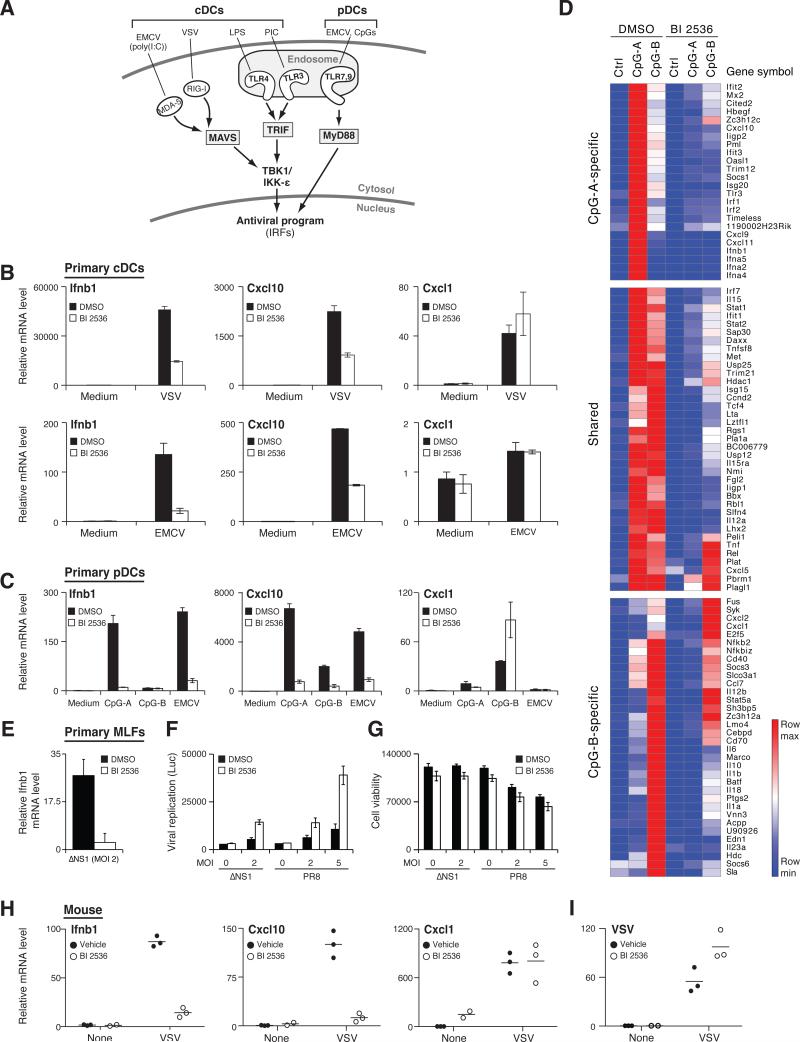
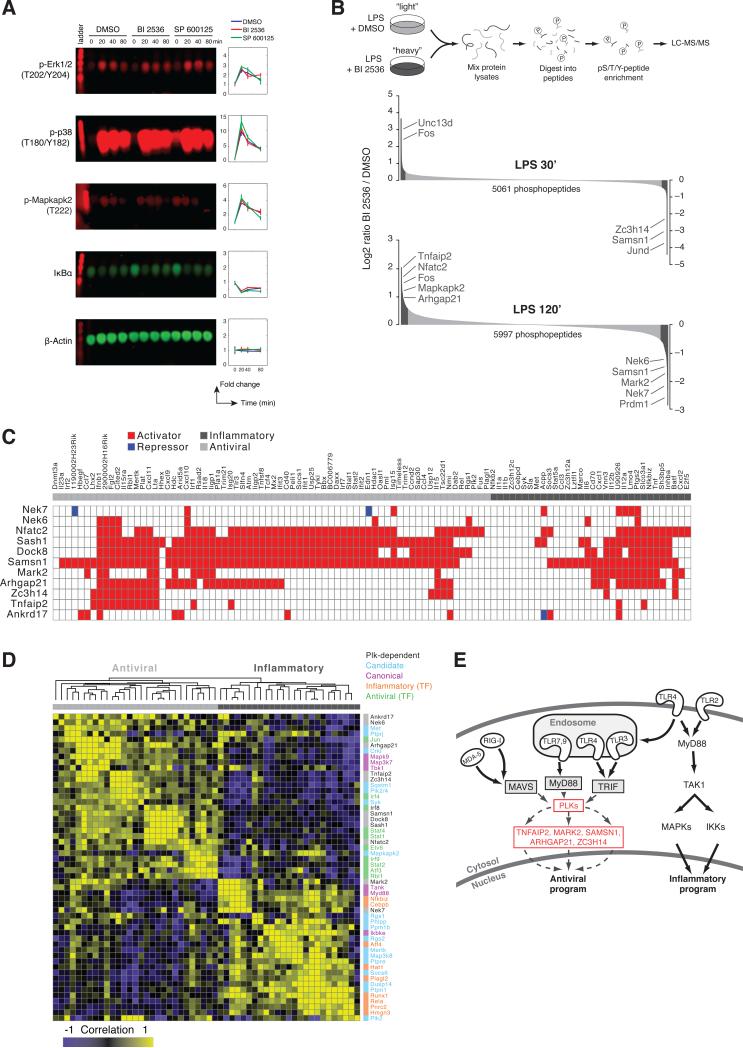
References
-
- Amit I, Citri A, Shay T, Lu Y, Katz M, Zhang F, Tarcic G, Siwak D, Lahad J, Jacob-Hirsch J, et al. A module of negative feedback regulators defines growth factor signaling. Nat Genet. 2007;39:503–512. - PubMed
-
- Archambault V, Glover DM. Polo-like kinases: conservation and divergence in their functions and regulation. Nat Rev Mol Cell Biol. 2009;10:265–275. - PubMed
-
- Banchereau J, Pascual V. Type I interferon in systemic lupus erythematosus and other autoimmune diseases. Immunity. 2006;25:383–392. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
