

Microbial domestication signatures of Lactococcus lactis can be reproduced by experimental evolution
- PMID: 22080491
- PMCID: PMC3246198
- DOI: 10.1101/gr.121285.111
Microbial domestication signatures of Lactococcus lactis can be reproduced by experimental evolution
Abstract
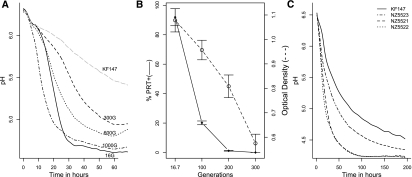
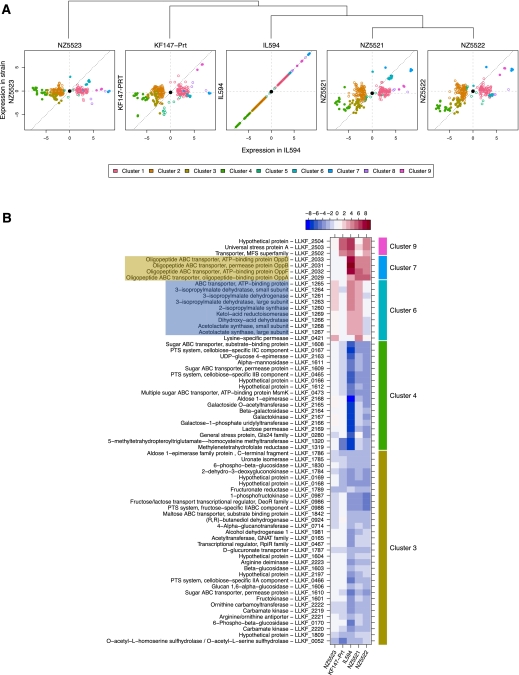
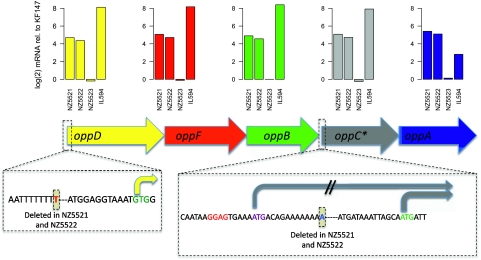
Experimental evolution is a powerful approach to unravel how selective forces shape microbial genotypes and phenotypes. To this date, the available examples focus on the adaptation to conditions specific to the laboratory. The lactic acid bacterium Lactococcus lactis naturally occurs on plants and in dairy environments, and it is proposed that dairy strains originate from the plant niche. Here we investigate the adaptation of a L. lactis strain isolated from a plant to a dairy niche by propagating it for 1000 generations in milk. Two out of three independently evolved strains displayed significantly increased acidification rates and biomass yields in milk. Genome resequencing, revealed six, seven, and 28 mutations in the three strains, including point mutations in loci related to amino acid biosynthesis and transport and in the gene encoding MutL, which is involved in DNA mismatch repair. Two strains lost a conjugative transposon containing genes important in the plant niche but dispensable in milk. A plasmid carrying an extracellular protease was introduced by transformation. Although improving growth rate and growth yield significantly, the plasmid was rapidly lost. Comparative transcriptome and phenotypic analyses confirmed that major physiological changes associated with improved growth in milk relate to nitrogen metabolism and the loss or down-regulation of several pathways involved in the utilization of complex plant polymers. Reproducing the transition from the plant to the dairy niche through experimental evolution revealed several genome, transcriptome, and phenotype signatures that resemble those seen in strains isolated from either niche.
Figures
References
-
- Bachmann H, de Wilt L, Kleerebezem M, van Hylckama Vlieg JE 2010. Time-resolved genetic responses of Lactococcus lactis to a dairy environment. Environ Microbiol 12: 1260–1270 - PubMed
-
- Barrick JE, Yu DS, Yoon SH, Jeong H, Oh TK, Schneider D, Lenski RE, Kim JF 2009. Genome evolution and adaptation in a long-term experiment with Escherichia coli. Nature 461: 1243–1247 - PubMed
-
- Beaumont HJ, Gallie J, Kost C, Ferguson GC, Rainey PB 2009. Experimental evolution of bet hedging. Nature 462: 90–93 - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources