

Review
doi: 10.1038/nrmicro2688.
The myriad roles of cyclic AMP in microbial pathogens: from signal to sword
Affiliations
- PMID: 22080930
- PMCID: PMC3785115
- DOI: 10.1038/nrmicro2688
Item in Clipboard
Review
The myriad roles of cyclic AMP in microbial pathogens: from signal to sword
Nat Rev Microbiol.
.
Abstract
All organisms must sense and respond to their external environments, and this signal transduction often involves second messengers such as cyclic nucleotides. One such nucleotide is cyclic AMP, a universal second messenger that is used by diverse forms of life, including mammals, fungi, protozoa and bacteria. In this review, we discuss the many roles of cAMP in bacterial, fungal and protozoan pathogens and its contributions to microbial pathogenesis. These roles include the coordination of intracellular processes, such as virulence gene expression, with extracellular signals from the environment, and the manipulation of host immunity by increasing cAMP levels in host cells during infection.
Figures
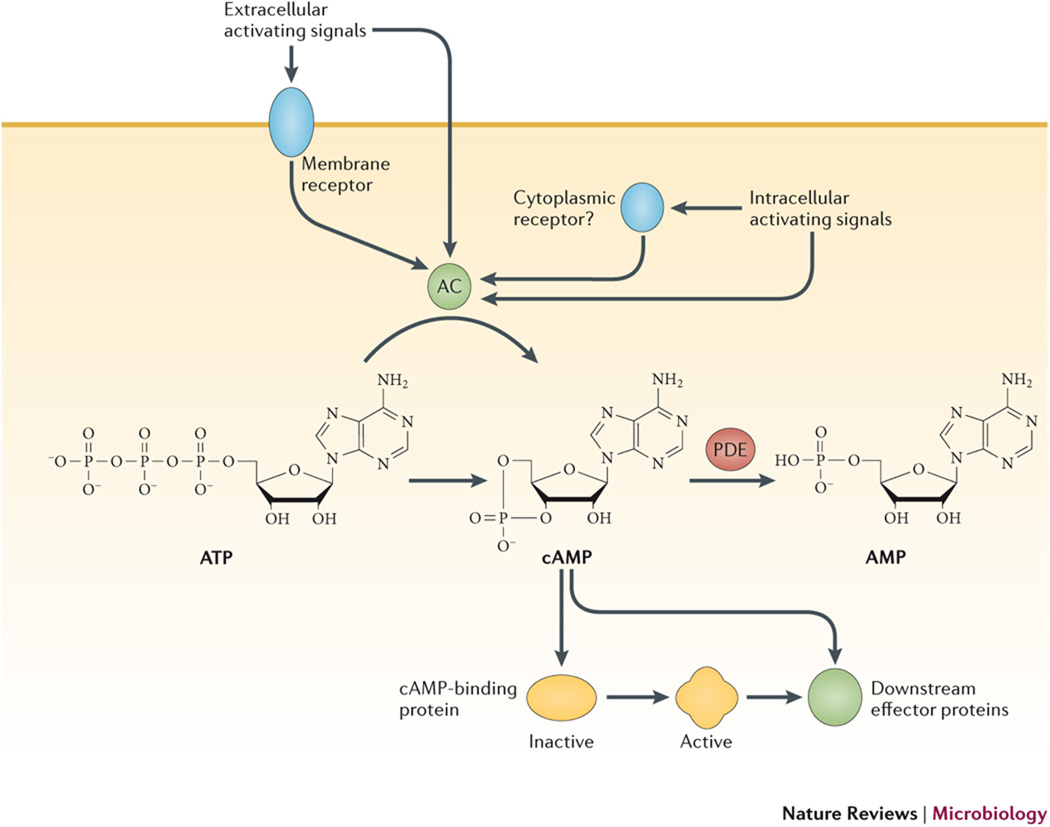
Adenylyl cyclases (ACs) can be directly or indirectly activated at the post-translational level by a number of environmental signals. Indirect activation pathways often involve membrane receptors that transmit extracellular signals to the AC by a phosphorylation event, although other cytoplasmic factors (“Factor X” in the figure) may also be required. cAMP then relays the signal to downstream effector proteins (DEP) either directly or indirectly by allosterically activating one or more cAMP-binding proteins. These cAMP-binding proteins may themselves be effector proteins, such as transcription factors or cyclic nucleotide gated channel (CNG) proteins, or they may be intermediaries, such as the regulatory subunits of PKA, which control activation of effector proteins further downstream. The inset shows the synthesis of cAMP by ACs, which catalyze conversion of ATP into cAMP and inorganic pyrophosphate while linking the remaining 5' phosphate with the 3' ribose carbon. cAMP is degraded by phosphodiesterases (PDEs) that catalyze the hydrolysis of the 3' linkage (marked with arrowhead), leaving adenosine 5' phosphate (not shown).
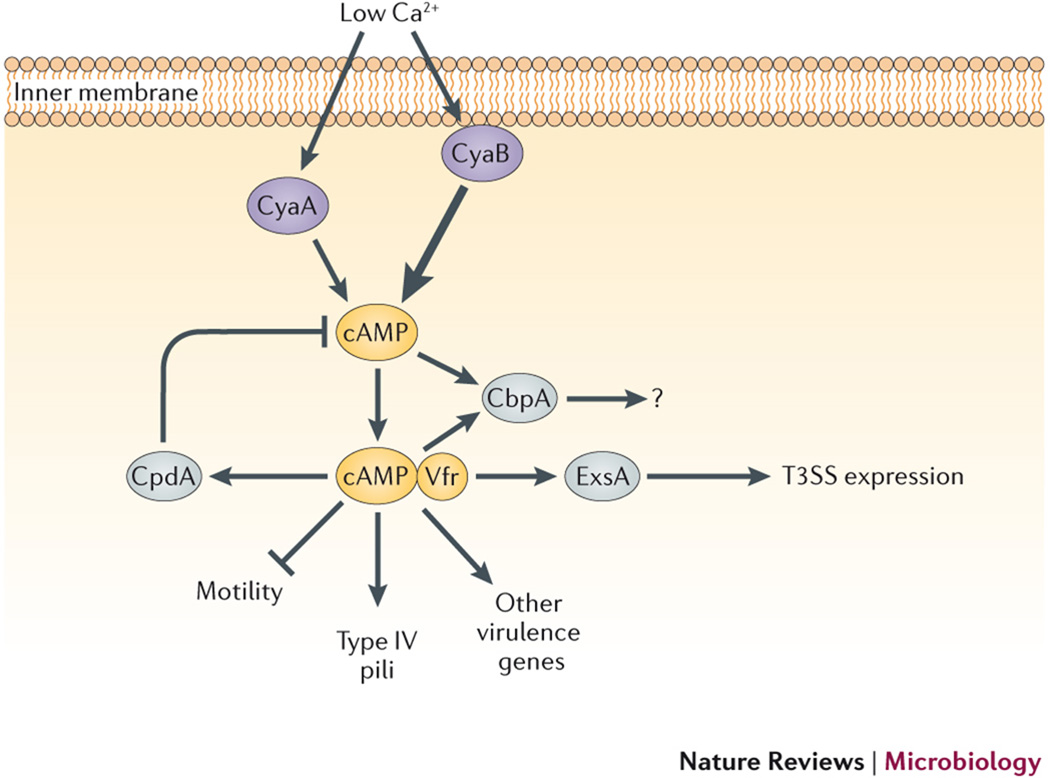
Two ACs in P. aeruginosa, CyaA and CyaB, are induced by low Ca2+ concentrations, although CyaB is the dominant producer of cAMP, as indicated by the thicker arrow. cAMP binding to the CRP family transcription factor, Vfr (virulence factor regulator), regulates expression of a broad range of phenotypes. Intermediate regulatory factors are noted, where known, including LasI (LuxI-type acyl homoserine lactone (AHL) synthase) and LasR (regulator of the Las quorum sensing system), as well as the transcription regulator ExsA (exoenzyme secretion protein A). Arrows indicate positive regulation, while blunted lines indicate repression.
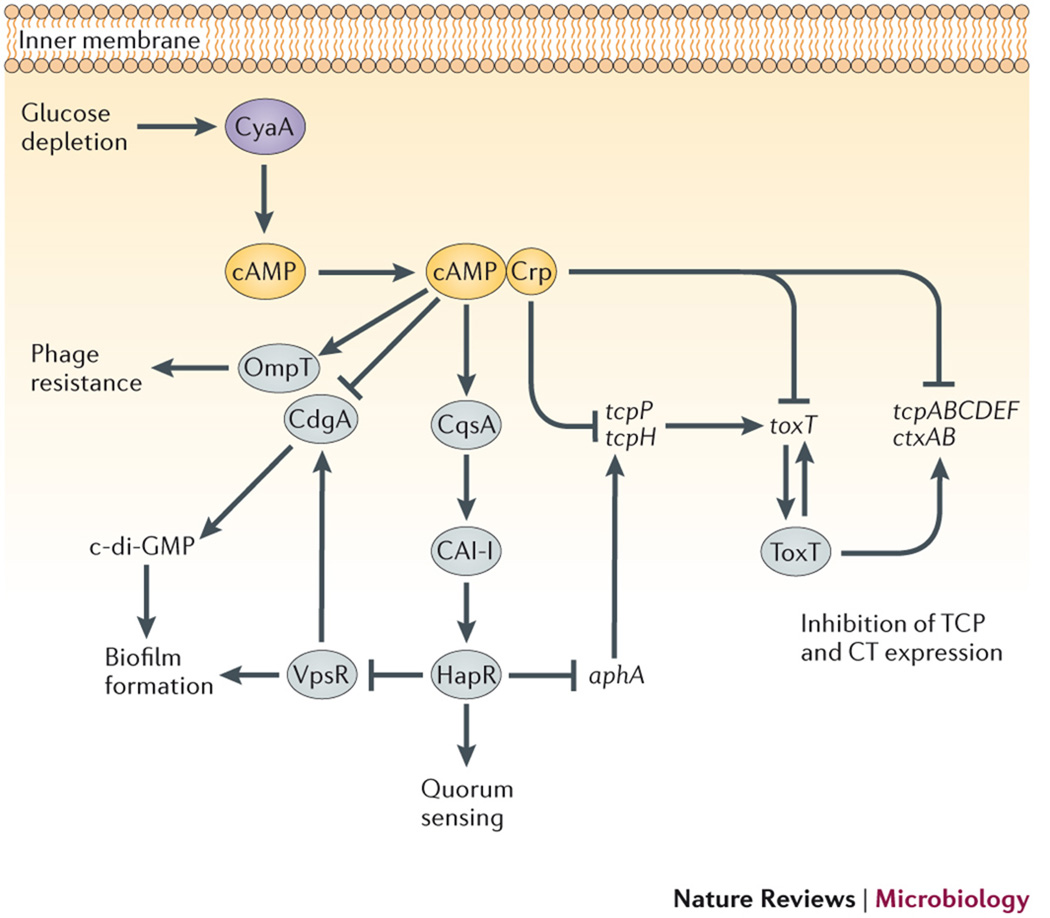
cAMP signaling integrates carbon source information with a wide range of pathogenesis-associated processes in V. cholerae, through its effects on gene regulation. Biofilm formation, quorum sensing, virulence gene expression and phage sensitivity are all regulated either directly or indirectly by cAMP-CRP. cAMP-CRP regulates several individual processes by multiple pathways, emphasizing the likely importance of cAMP signaling throughout the infection cycle. Gene names: ompT, outer membrane protein T; cqsA, CAI-1 synthase ; vpsR, Vibrio polysaccharide regulator; ahpAB, transcriptional regulators; toxT, toxin regulator T; ctxAB, cholera toxin A and B (operon); tcpA-F, toxin-co-regulated pilus genes A to F (operon). Protein names: CyaA, adenylyl cyclase A; CrpVc, cAMP receptor protein; ToxR, toxin regulator R; CdgA, cyclic diguanylate cyclase A; HapR, hemagglutinin protease regulator; ToxT, toxin regulator T. Small molecules: di-cGMP, di-cyclic GMP; CAI-I, cholera autoinducer 1; HCO3−, bicarbonate ion.

cAMP signaling in M. tuberculosis is complex owing to its large number of adenylyl cyclases (ACs). The 'Rv' numbers listed in top section without boxes refer to the names of ten individual M. tuberculosis ACs that have demonstrated activity. Six additional putative ACs are not shown. Green boxes (top) include signals that activate the specific ACs indicated by arrows. Crp, Cmr and Rv0998 are cAMP-dependent effector proteins, which are activated by association with cAMP. However, a direct binding of cAMP to Cmr has not been demonstrated. CrpMt and Cmr are transcription factors, while Rv0998 is a protein acetylase. The other 'Rv' numbers in yellow boxes (middle section) correspond to seven other putative cAMP binding proteins. Blue boxes indicate the respective functions of specific cAMP binding proteins. Secretion of cAMP into macrophages is indicated by an arrow that connects cAMP (middle) to a blue box. Rv0805 is a phosphodiesterase that has low activity on cAMP. Arrows indicate positive activation, while the blunt ended line indicates degradation.

Bacterial pathogens use numerous strategies to elevate cAMP levels within host cells during infection. Shown are the ADP-ribosylating exotoxins that increase the activity of endogenous host ACs (top, AC Modulating Toxins); the exported host-activated bacterial ACs produced by Bordetella pertussis (CyaA), Bacillus anthracis (EF), Pseudomonas aeruginosa (ExoY) and Yersinia pestis (Yp AC), (middle, AC Toxins); the activation of host ACs through coordinate signaling of complement C5a receptor C5aR (pink) and Toll-like receptor TLR2 (orange) stimulated by Porphyromonas gingivalis; and the direct secretion of cAMP by Mycobacterium tuberculosis (bottom). Red triangles represent calmodulin, red squares represent an unknown host activation factor, a purple diamond is C5a and the yellow diamonds are cAMP. Gαs and Gαi are stimulatory and inhitory subunits of heterotrimeric G proteins, respectively.

Both Cryptococcus neoformans (left) and Candida albicans (right) use their highly conserved cAMP/PKA pathways to regulate expression of key virulence determinants. However, the functions of these pathways differ. Matching shapes and colors are used to indicate orthologs in both fungi. Arrows denote positive regulatory effects while flat ended lines indicate inhibitory effects. Protein abbreviations for C. neoformans: Gpr4, G protein receptor; Gpa1, G protein G alpha subunit; Crg2, regulator of G protein signaling; Pkr1, PKA regulatory subunit; Cac1, adenylyl cyclase; Pka1, PKA catalytic subunit; Nrg1, transcription factor ; Can2, carbonic anhydrase; Ras1, guanosine nucleotide binding protein; Aca1, AC-associated protein; Pde1, phosphodiesterase. Protein abbreviations for C. albicans: Gpr1, G protein-coupled receptor; Gpa2, G protein alpha subunit; GAP, GTPase activator protein; Cyr1, adenylyl cyclase; Tpk1 and Tpk2, protein kinases; Nce103, carbonic anhydrase; Ras1, guanosine nucleotide binding protein; Cap1, cyclase associated protein; Pde2, phosphodiesterase; Bcy1, PKA regulatory subunit; Efg1 and Flo8, transcription factors.

ACα contains a domain with a putative K+ channel in Plasmodium. Extracellular K+ influx (1) activates AC activity, resulting in the increase of cAMP (2). Elevation of cAMP levels activates PKA (3) and leads to exocytosis of apical granules (micronemes) (4) that carry transmembrane proteins, which are necessary for invasion of host hepatocytes by Plasmodium sporozoites.
References
-
- Beavo JA, Brunton LL. Cyclic nucleotide research -- still expanding after half a century. Nat Rev Mol Cell Biol. 2002;3:710–718. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
