

A MEK-independent role for CRAF in mitosis and tumor progression
- PMID: 22081024
- PMCID: PMC3233644
- DOI: 10.1038/nm.2464
A MEK-independent role for CRAF in mitosis and tumor progression
Abstract
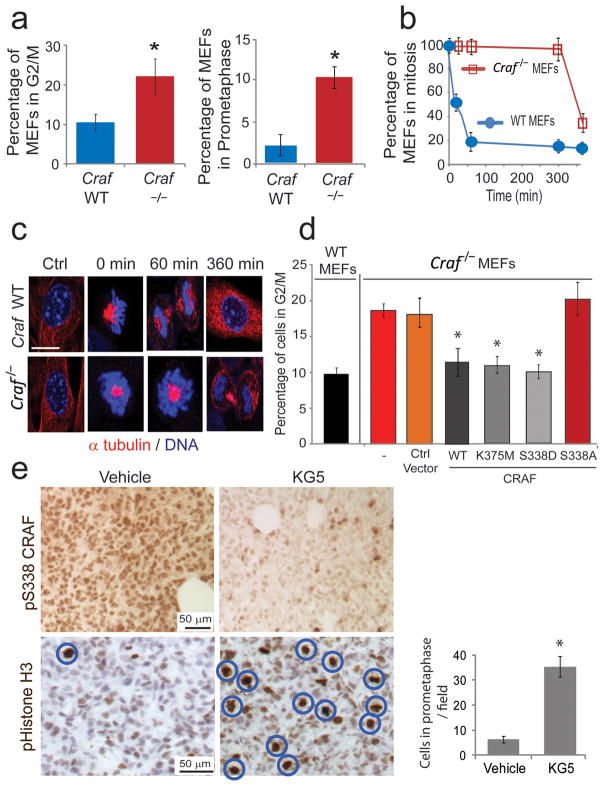
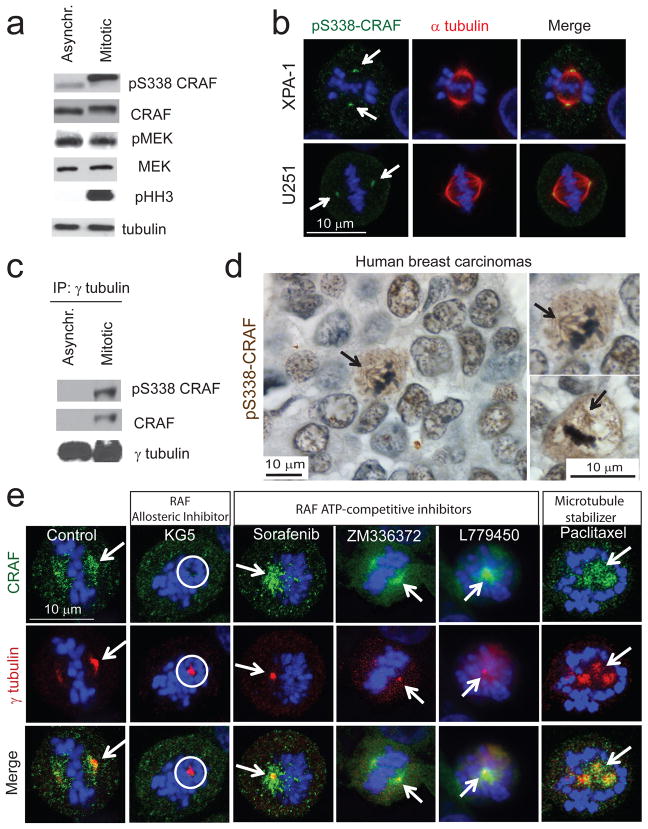
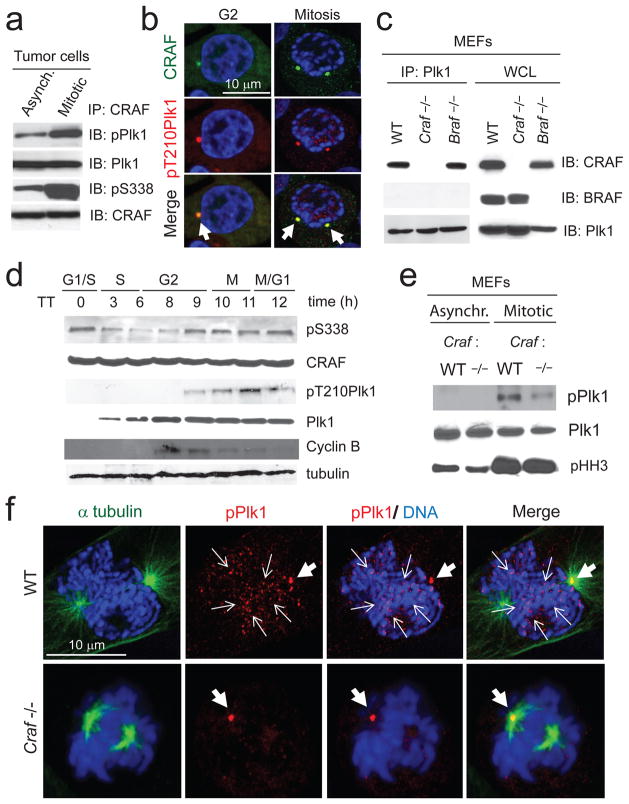
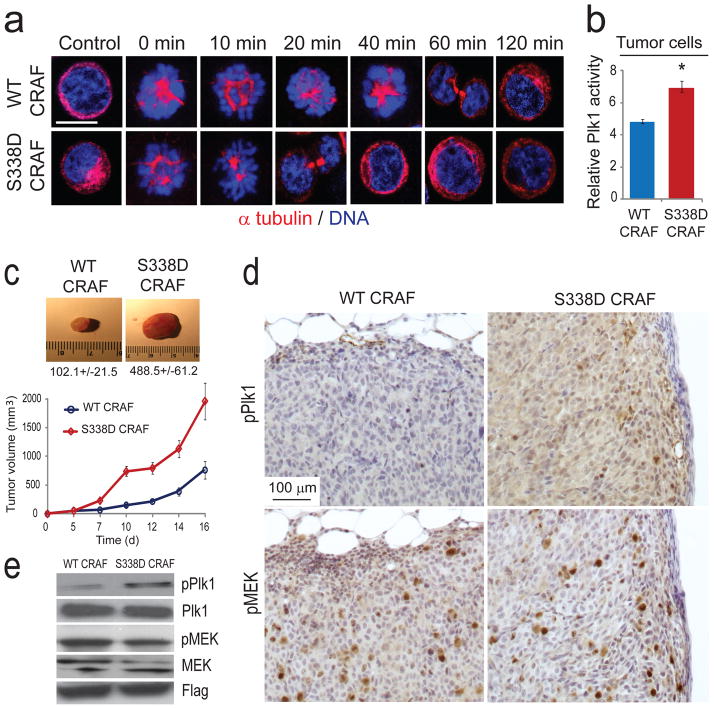
RAF kinases regulate cell proliferation and survival and can be dysregulated in tumors. The role of RAF in cell proliferation has been linked to its ability to activate mitogen-activated protein kinase kinase 1 (MEK) and mitogen-activated protein kinase 1 (ERK). Here we identify a MEK-independent role for RAF in tumor growth. Specifically, in mitotic cells, CRAF becomes phosphorylated on Ser338 and localizes to the mitotic spindle of proliferating tumor cells in vitro as well as in murine tumor models and in biopsies from individuals with cancer. Treatment of tumors with allosteric inhibitors, but not ATP-competitive RAF inhibitors, prevents CRAF phosphorylation on Ser338 and localization to the mitotic spindle and causes cell-cycle arrest at prometaphase. Furthermore, we identify phospho-Ser338 CRAF as a potential biomarker for tumor progression and a surrogate marker for allosteric RAF blockade. Mechanistically, CRAF, but not BRAF, associates with Aurora kinase A (Aurora-A) and Polo-like kinase 1 (Plk1) at the centrosomes and spindle poles during G2/M. Indeed, allosteric or genetic inhibition of phospho-Ser338 CRAF impairs Plk1 activation and accumulation at the kinetochores, causing prometaphase arrest, whereas a phospho-mimetic Ser338D CRAF mutant potentiates Plk1 activation, mitosis and tumor progression in mice. These findings show a previously undefined role for RAF in tumor progression beyond the RAF-MEK-ERK paradigm, opening new avenues for targeting RAF in cancer.
Conflict of interest statement
The authors declare no competing financial interests. Published online at
Reprints and permissions information is available online at
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
