

Synthetic biology approach to reconstituting the ubiquitylation cascade in bacteria
- PMID: 22081111
- PMCID: PMC3261559
- DOI: 10.1038/emboj.2011.397
Synthetic biology approach to reconstituting the ubiquitylation cascade in bacteria
Abstract
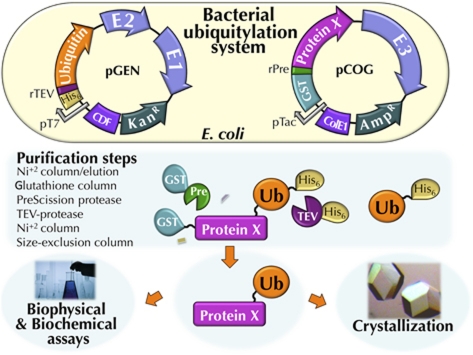
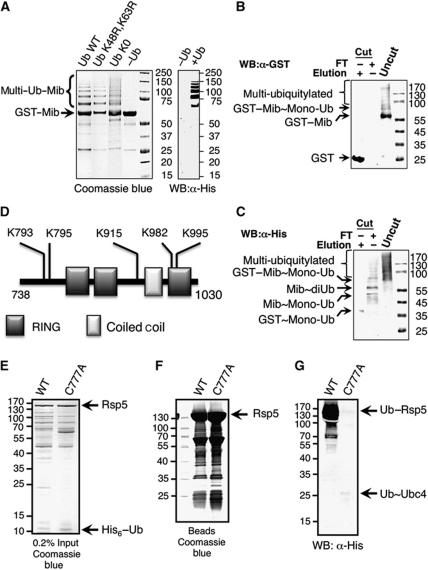
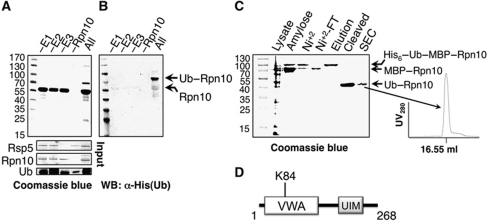
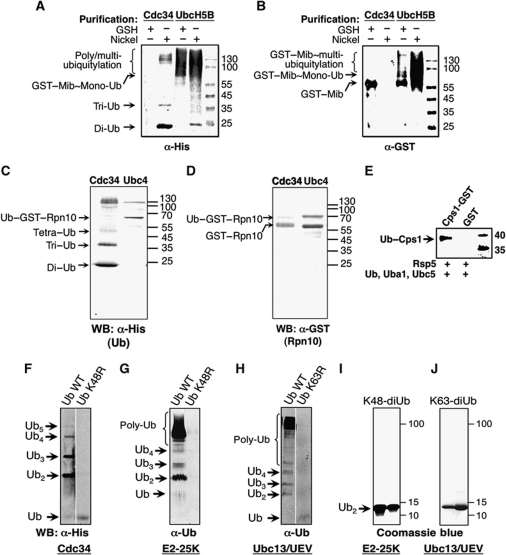
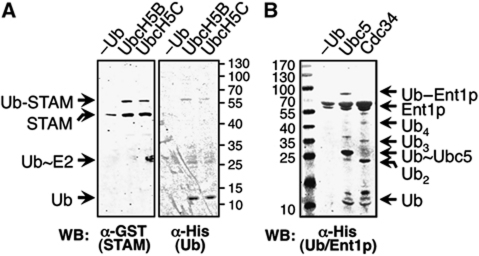
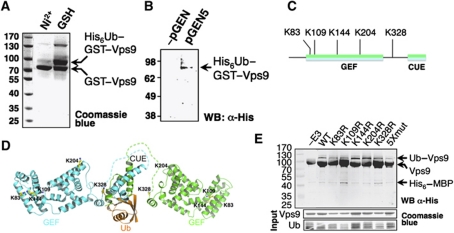
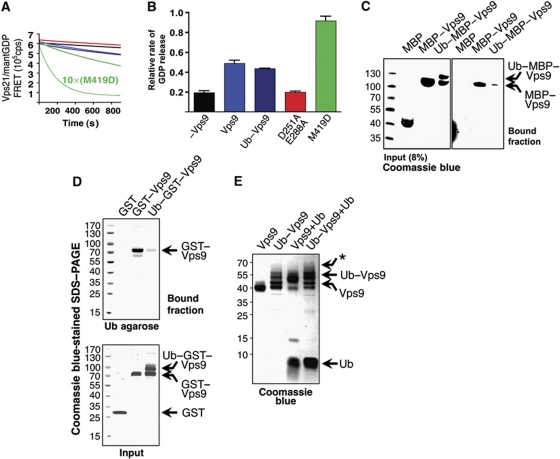
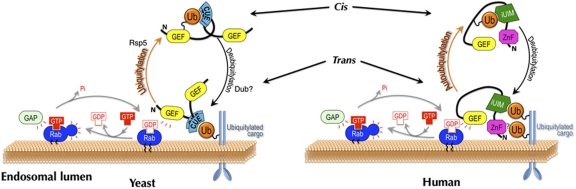
Covalent modification of proteins with ubiquitin (Ub) is widely implicated in the control of protein function and fate. Over 100 deubiquitylating enzymes rapidly reverse this modification, posing challenges to the biochemical and biophysical characterization of ubiquitylated proteins. We circumvented this limitation with a synthetic biology approach of reconstructing the entire eukaryotic Ub cascade in bacteria. Co-expression of affinity-tagged substrates and Ub with E1, E2 and E3 enzymes allows efficient purification of ubiquitylated proteins in milligram quantity. Contrary to in-vitro assays that lead to spurious modification of several lysine residues of Rpn10 (regulatory proteasomal non-ATPase subunit), the reconstituted system faithfully recapitulates its monoubiquitylation on lysine 84 that is observed in vivo. Mass spectrometry revealed the ubiquitylation sites on the Mind bomb E3 ligase and the Ub receptors Rpn10 and Vps9. Förster resonance energy transfer (FRET) analyses of ubiquitylated Vps9 purified from bacteria revealed that although ubiquitylation occurs on the Vps9-GEF domain, it does not affect the guanine nucleotide exchanging factor (GEF) activity in vitro. Finally, we demonstrated that ubiquitylated Vps9 assumes a closed structure, which blocks additional Ub binding. Characterization of several ubiquitylated proteins demonstrated the integrity, specificity and fidelity of the system, and revealed new biological findings.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Azem A, Weiss C, Goloubinoff P (1998) Structural analysis of GroE chaperonin complexes using chemical cross-linking. Methods Enzymol 290: 253–268 - PubMed
-
- Baba D, Maita N, Jee JG, Uchimura Y, Saitoh H, Sugasawa K, Hanaoka F, Tochio H, Hiroaki H, Shirakawa M (2005) Crystal structure of thymine DNA glycosylase conjugated to SUMO-1. Nature 435: 979–982 - PubMed
-
- Bachmair A, Finley D, Varshavsky A (1986) In vivo half-life of a protein is a function of its amino-terminal residue. Science 234: 179–186 - PubMed
-
- Chatterjee C, McGinty RK, Fierz B, Muir TW (2010) Disulfide-directed histone ubiquitylation reveals plasticity in hDot1L activation. Nat Chem Biol 6: 267–269 - PubMed
-
- Chen J, Ai Y, Wang J, Haracska L, Zhuang Z (2010) Chemically ubiquitylated PCNA as a probe for eukaryotic translesion DNA synthesis. Nat Chem Biol 6: 270–272 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
