

Suppression of antigen-specific CD4+ T cell activation by SRA/CD204 through reducing the immunostimulatory capability of antigen-presenting cell
- PMID: 22083206
- PMCID: PMC3288685
- DOI: 10.1007/s00109-011-0828-1
Suppression of antigen-specific CD4+ T cell activation by SRA/CD204 through reducing the immunostimulatory capability of antigen-presenting cell
Abstract
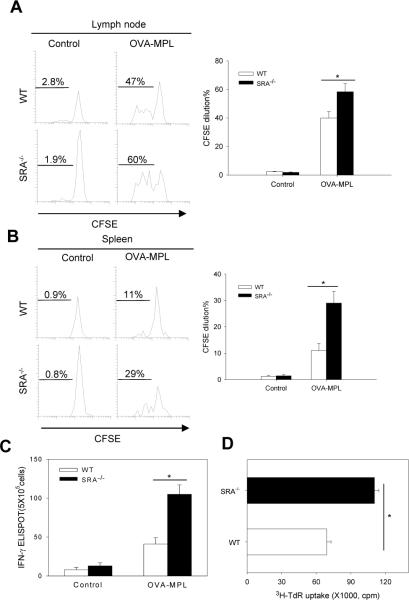
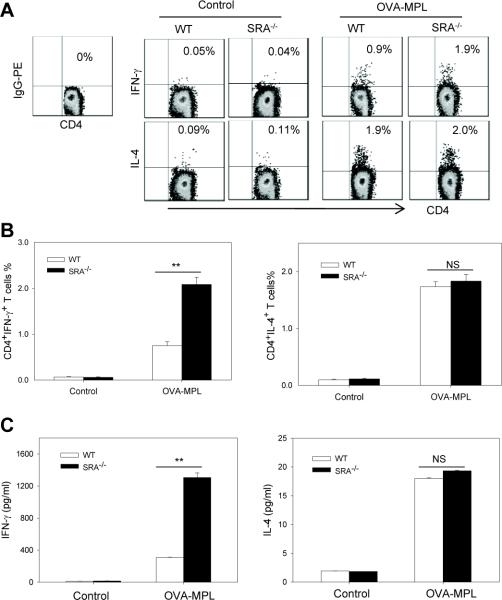
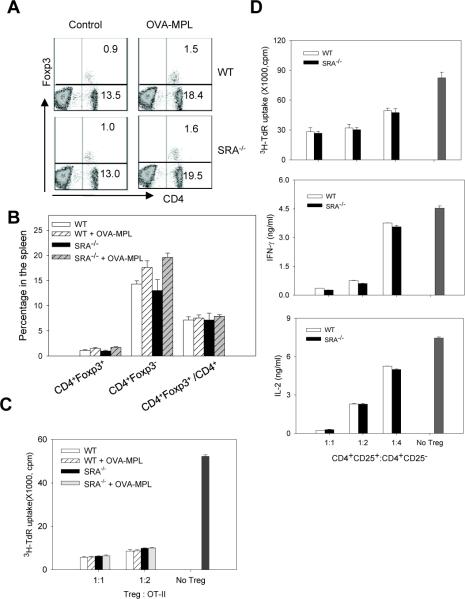
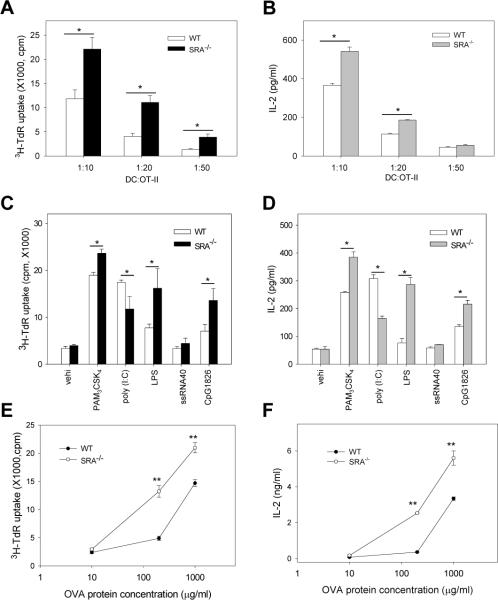
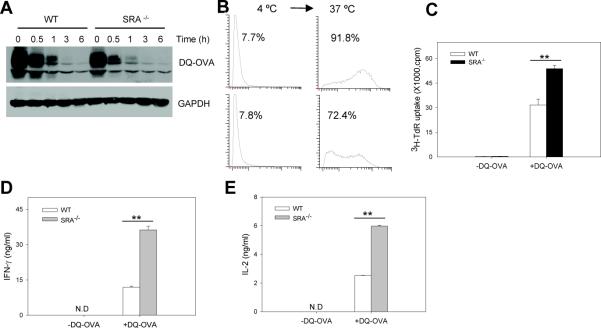
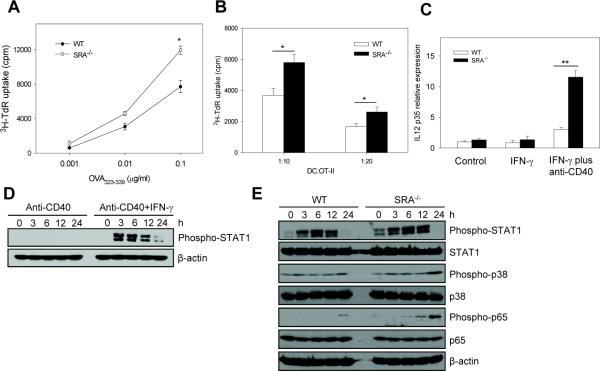
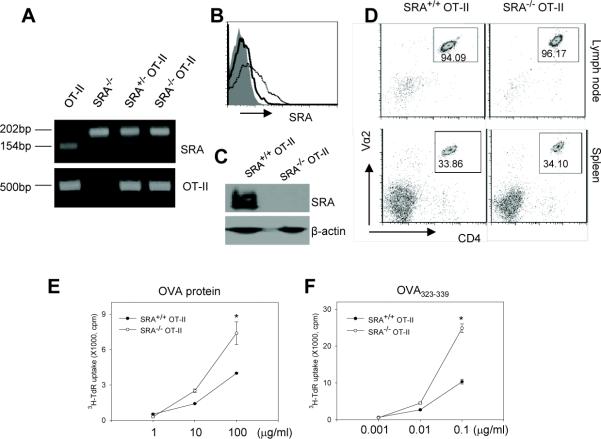
Pattern recognition scavenger receptor SRA/CD204, primarily expressed on specialized antigen-presenting cells (APCs), including dendritic cells (DCs) and macrophages, has been implicated in multiple physiological and pathological processes, including atherosclerosis, Alzheimer's disease, endotoxic shock, host defense, and cancer development. SRA/CD204 was also recently shown to function as an attenuator of vaccine response and antitumor immunity. Here, we, for the first time, report that SRA/CD204 knockout (SRA(-/-)) mice developed a more robust CD4(+) T cell response than wild-type mice after ovalbumin immunization. Splenic DCs from the immunized SRA(-/-) mice were much more efficient than those from WT mice in stimulating naïve OT-II cells, indicating that the suppressive activity of SRA/CD204 is mediated by DCs. Strikingly, antigen-exposed SRA(-/-) DCs with or without lipopolysaccharide treatment exhibited increased T-cell-stimulating activity in vitro, which was independent of the classical endocytic property of the SRA/CD204. Additionally, absence of SRA/CD204 resulted in significantly elevated IL12p35 expression in DCs upon CD40 ligation plus interferon gamma (IFN-γ) stimulation. Molecular studies reveal that SRA/CD204 inhibited the activation of STAT1, mitogen activated protein kinase p38, and nuclear factor-kappa B signaling activation in DCs treated with anti-CD40 antibodies and IFN-γ. Furthermore, splenocytes from the generated SRA(-/-) OT-II mice showed heightened proliferation upon stimulation with OVA protein or MHC-II-restricted OVA(323-339) peptide compared with cells from the SRA(+/+) OT-II mice. These results not only establish a new role of SRA/CD204 in limiting the intrinsic immunogenicity of APCs and CD4(+) T cell activation but also provide additional insights into the molecular mechanisms involved in the immune suppression by this molecule.
Figures
References
-
- Suzuki H, Kurihara Y, Takeya M, Kamada N, Kataoka M, Jishage K, Ueda O, Sakaguchi H, Higashi T, Suzuki T, Takashima Y, Kawabe Y, Cynshi O, Wada Y, Honda M, Kurihara H, Aburatani H, Doi T, Matsumoto A, Azuma S, Noda T, Toyoda Y, Itakura H, Yazaki Y, Steinbrecher UP, Ishinash S, Maeda N, Gordon S, Kodama T. A role for macrophage scavenger receptors in atherosclerosis and susceptibility to infection. Nature. 1997;386:292–296. - PubMed
-
- Peiser L, De Winther MP, Makepeace K, Hollinshead M, Coull P, Plested J, Kodama T, Moxon ER, Gordon S. The class A macrophage scavenger receptor is a major pattern recognition receptor for Neisseria meningitidis which is independent of lipopolysaccharide and not required for secretory responses. Infect Immun. 2002;70:5346–5354. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
