

Cell cycle adaptations of embryonic stem cells
- PMID: 22084091
- PMCID: PMC3228440
- DOI: 10.1073/pnas.1116794108
Cell cycle adaptations of embryonic stem cells
Abstract
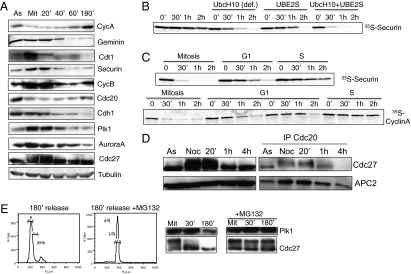
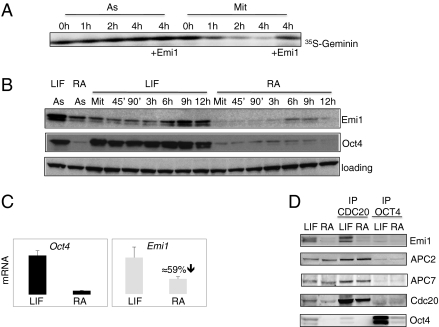
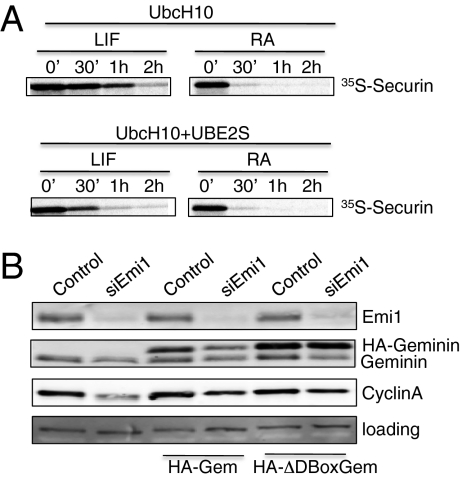
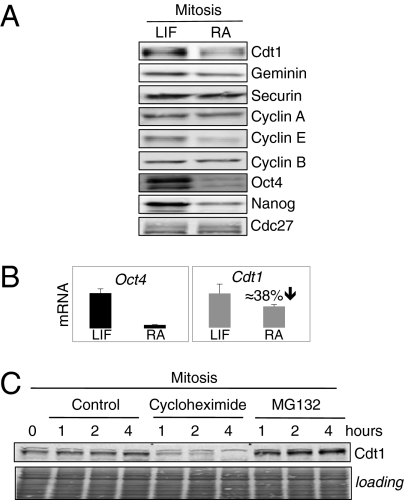
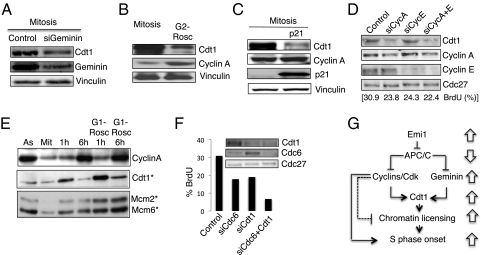
ES cells proliferate with very short gap phases yet maintain their capacity to differentiate. It had been thought that the levels of cyclins and other substrates of ubiquitin ligase APC/C remain nearly constant and Cdk activity remains constitutively high in mouse ES cells. Here we demonstrate that APC/C (anaphase-promoting complex/cyclosome) enzyme is active in ES cells but attenuated by high levels of the Emi1 (early mitotic inhibitor-1) protein. Despite the presence of high Cdk activity during the G1 phase, chromatin can be effectively licensed for DNA replication and fast entry into the S phase can still occur. High Cdk activity during S-G2-M phases produces high levels of the DNA replication factor Cdt1, and this leads to efficient Mcm proteins loading on chromatin after mitotic exit. Although disturbing the usual balance between Cdk activity and APC/C activity found in somatic cells, a few key adaptations allow normal progression of a very rapid cell cycle.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Savatier P, Lapillonne H, Jirmanova L, Vitelli L, Samarut J. Analysis of the cell cycle in mouse embryonic stem cells. Methods Mol Biol. 2002;185:27–33. - PubMed
-
- Fujii-Yamamoto H, Kim JM, Arai K, Masai H. Cell cycle and developmental regulations of replication factors in mouse embryonic stem cells. J Biol Chem. 2005;280:12976–12987. - PubMed
-
- Bloom J, Cross FR. Multiple levels of cyclin specificity in cell-cycle control. Nat Rev Mol Cell Biol. 2007;8:149–160. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
