

Nutrient limitations alter cell division control and chromosome segregation through growth-related kinases and phosphatases
- PMID: 22084378
- PMCID: PMC3203466
- DOI: 10.1098/rstb.2011.0124
Nutrient limitations alter cell division control and chromosome segregation through growth-related kinases and phosphatases
Abstract
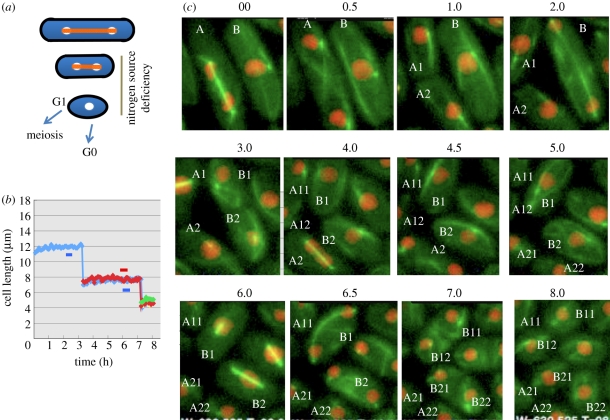
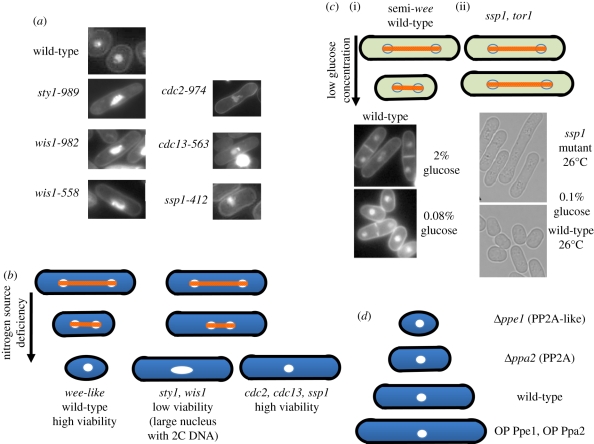
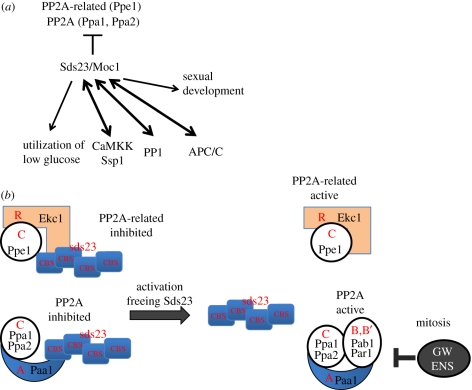
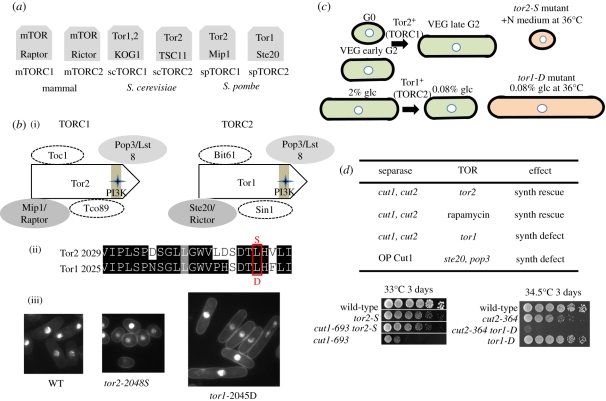
In dividing fission yeast Schizosaccharomyces pombe cells, the balance between Wee1 kinase and Cdc25 phosphatase which control the cyclin-dependent kinase (CDK) at the G2-M transition determines the rod-shaped cell length. Under nitrogen source starvation or glucose limitation, however, cell size determination is considerably modulated, and cell size shortening occurs for wild-type cells. For several mutants of kinases or phosphatases, including CDK, target of rapamycin complex (TORC) 1 and 2, stress-responsive mitogen-activated protein kinase (MAPK) Sty1/Spc1, MAPK kinase Wis1, calcium- and calmodulin-dependent protein kinase kinase-like Ssp1, and type 2A and 2A-related phosphatases inhibitor Sds23, this cell shortening does not normally occur. In tor1 and ssp1 mutants, cell elongation is observed. Sds23 that binds to and inhibits 2A and 2A-related phosphatases is synergistic with Ssp1 in the cell size determination and survival under low glucose and nitrogen source. Tor2 (TORC1) is required for growth, whereas Tor1 (TORC2) is needed for determining division size according to different nutrient conditions. Surprisingly, in growth-diminished tor2 mutant or rapamycin-treated cells, the requirement of separase/Cut1-securin/Cut2 essential for chromosome segregation is greatly alleviated. By contrast, defects of tor1 with secruin/cut2 or overproduction of Cut1 are additive. While Tor1 and Tor2 are opposite in their apparent functions, both may actually coordinate cell division with growth in response to the changes in nutrients.
Figures
References
-
- Mitchison J. 1971. The biology of the cell cycle. London, UK: Cambridge University Press.
-
- Mitchison J. M. 1957. The growth of single cells. I. Schizosaccharomyces pombe. Exp. Cell Res. 13, 244–26210.1016/0014-4827(57)90005-8 (doi:10.1016/0014-4827(57)90005-8) - DOI - DOI - PubMed
-
- Nurse P., Thuriaux P., Nasmyth K. 1976. Genetic control of the cell division cycle in the fission yeast Schizosaccharomyces pombe. Mol. Gen. Genet. 146, 167–17810.1007/BF00268085 (doi:10.1007/BF00268085) - DOI - DOI - PubMed
-
- Thuriaux P., Nurse P., Carter B. 1978. Mutants altered in the control co-ordinating cell division with cell growth in the fission yeast Schizosaccharomyces pombe. Mol. Gen. Genet. 161, 215–22010.1007/BF00274190 (doi:10.1007/BF00274190) - DOI - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials