

Review
doi: 10.1534/genetics.111.127019.
Transcriptional regulation in Saccharomyces cerevisiae: transcription factor regulation and function, mechanisms of initiation, and roles of activators and coactivators
Affiliations
- PMID: 22084422
- PMCID: PMC3213380
- DOI: 10.1534/genetics.111.127019
Item in Clipboard
Review
Transcriptional regulation in Saccharomyces cerevisiae: transcription factor regulation and function, mechanisms of initiation, and roles of activators and coactivators
Genetics.
2011 Nov.
Abstract
Here we review recent advances in understanding the regulation of mRNA synthesis in Saccharomyces cerevisiae. Many fundamental gene regulatory mechanisms have been conserved in all eukaryotes, and budding yeast has been at the forefront in the discovery and dissection of these conserved mechanisms. Topics covered include upstream activation sequence and promoter structure, transcription factor classification, and examples of regulated transcription factor activity. We also examine advances in understanding the RNA polymerase II transcription machinery, conserved coactivator complexes, transcription activation domains, and the cooperation of these factors in gene regulatory mechanisms.
Figures

DNA-binding domains of five yeast transcription factors. Blue spheres, Zn; gray spheres, DNA. Protein data bank (http://www.biochem.ucl.ac.uk/bsm/prot_dna/prot_dna.html ) accession codes are in parentheses. (A) C2H2 zinc fingers of Adr1 (2ADR); (B) C6 (zinc knuckle) of Gal4 (1D66); (C) bZIP structure of Gcn4 (1YSA); (D) bHLH of Pho4 (1AOA); (E) helix-turn-helix of Matα2 and winged helix of Mcm1 (1MNM).
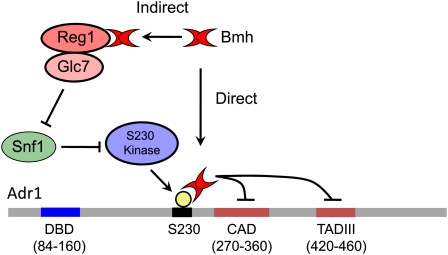
Model for Bmh regulation of Adr1 activity that could explain the direct and indirect roles of Bmh in the repression of Adr1-dependent gene expression. The direct role of Bmh involves binding to the S230-phosphorylated regulatory domain and inhibition of Adr1 ADs. Snf1 is implicated in reversing this direct inhibition because it is required to promote the dephosphorylation of S230 and thus inhibit Bmh binding. Snf1 is also involved in the indirect role of Bmh because Snf1 is inactivated by the Reg1–Glc7 complex in which Bmh plays an unknown role.
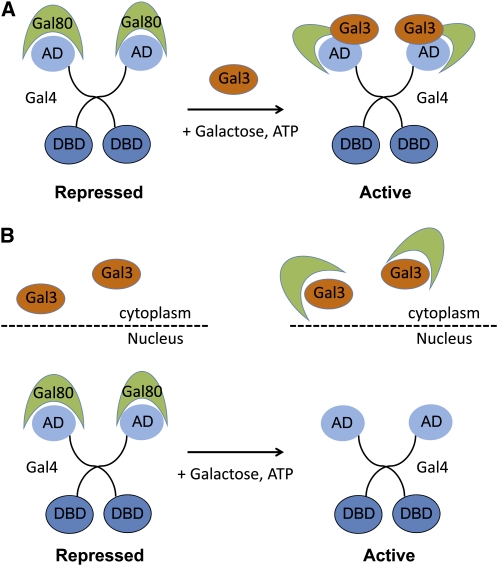
Two models for regulation of Gal4 by Gal80 and Gal3. (A) In the nondissociation model (Wightman et al. 2008), Gal80 may remain bound to DNA-bound Gal4 in the nucleus when it interacts with Gal3 and a structural change allows the Gal4 AD to interact with coactivators. (B) The dissociation model (Peng and Hopper 2000, suggests that Gal3 bound to galactose and ATP in the cytoplasm sequesters Gal80 from the nucleus and thus frees Gal4 AD for interaction with coactivators.
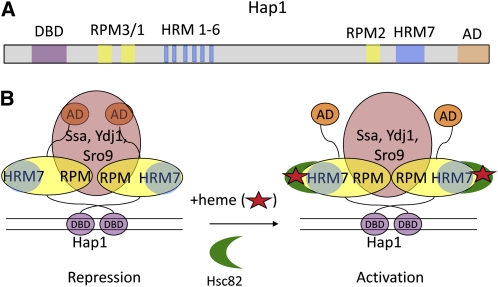
Model for the regulation of Hap1 by heme and molecular chaperones. (A) The diagram of the domain structure of Hap1 shows the DNA-binding domain (DBD, purple), activation domain (AD, tan), repression modules (RPM3/1, yellow), and heme-responsive motifs (HRM1-6, blue) that bind heme and recruit Hsp90 (Hsc82, green crescent in B) to activate transcription. (B) Model for the repression and activation of Hap1. In the absence of heme (red star), Hap1 is held in an inactive conformation by Ssa proteins (Hsp70) and co-chaperones Ydj1 and Sro9 (orange) that bind to repression modules (RPM). Activation occurs when heme binds to heme-responsive motifs (particularly HRM7), causing association with Hsc82 (Hsp90, green crescent), apparently without dissociation of the repressive chaperones (Lee and Zhang 2009).
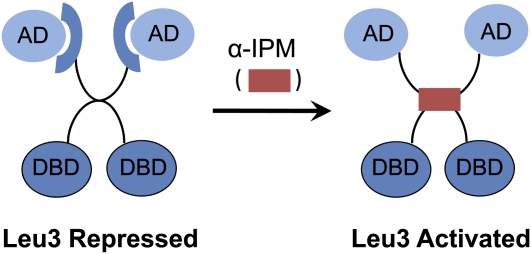
Leu3 regulation by intramolecular masking of the activation domain. In the absence of α-isopropylmalate (α-IPM), the inducing metabolite produced during leucine biosynthesis, Leu3 acts as a repressor. When α-IPM is present, Leu3 becomes an activator. Mutations in different parts of the central region either can make Leu3 a constitutive activator or inhibit Leu3 activity independently of α-−IPM. These mutations and the ability of Leu3 to be regulated by α-IPM in mammalian cell-free extracts suggest that its activation does not require another protein (Kohlhaw 2003).
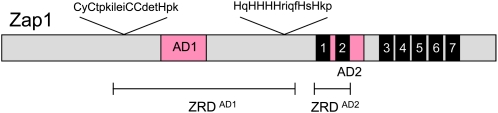
Zap1 regulation is mediated by Zn+2. The DNA-binding domain consists of C2H2 zinc fingers 3–7 (black boxes). AD1 is within the zinc-responsive domain (ZRD) that is Cys and His rich (Herbig et al. 2005). AD2 is within finger 2 but both fingers 1 and 2 are required for zinc responsiveness (Bird et al. 2000).
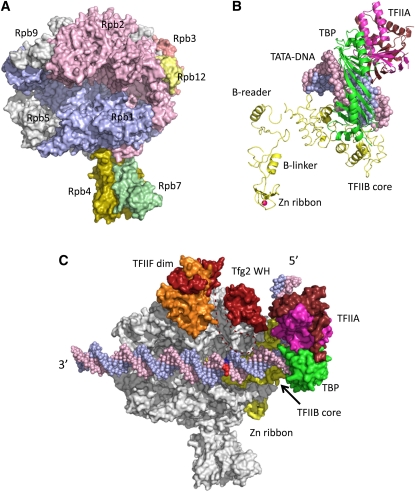
(A) RNA Pol II structure. Selected subunits are labeled. (B) Model for arrangement of TBP-TFIIB-TFIIA and TATA-DNA in the PIC. Red sphere, Zn. (C) Model for structure of the yeast Pol II PIC. The DNA template and nontemplate strands are blue and pink, and the base pair where DNA melting initiates is in dark blue and red; the TFIIF large and small subunits (Tfg1 and Tfg2) are orange and red; the TBP conserved domain (TBP) is green; TFIIB is yellow; and the large and small TFIIA subunits (Toa1 and Toa2) are brown and magenta. TFIIF dim, TFIIF dimerization domain; Tfg2 WH, Tfg2 winged helix domain; Zn ribbon, TFIIB N-terminal Zn ribbon domain. Dashed line represents a Tfg2 loop connecting the dimerization and winged helix domains. The 5′ and 3′ ends of the noncoding strand of promoter DNA are indicated.
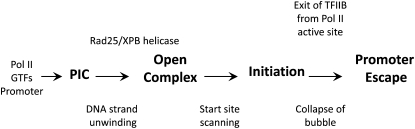
Steps in the pathway of yeast Pol II transcription initiation.
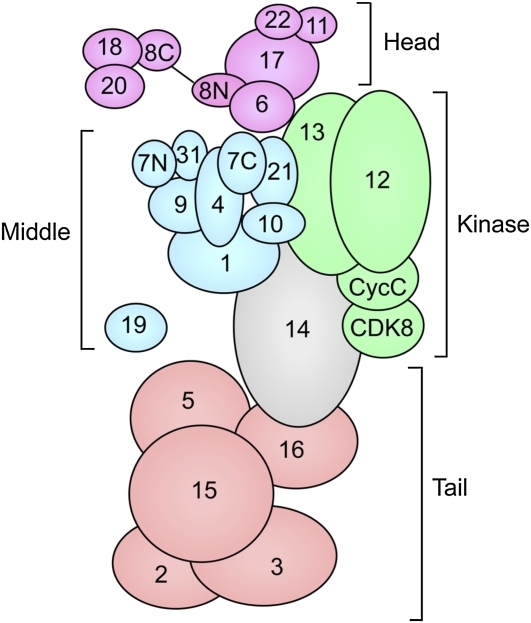
Organization of yeast Mediator and Mediator modules. Numbers indicate the Mediator subunit name. Head module, purple; middle module, blue; kinase module, green; tail module, orange. Med14 (Rgr1), in gray, is at the interface of the middle, kinase, and tail modules. Adapted from Koschubs, T., K. Lorenzen, S. Baumli, S. Sandstrom, A. J. Heck et al., 2010, Preparation and topology of the Mediator middle module. Nucleic Acids Res. 38: 3186–3195; by permission of Oxford University Press.

Organization of yeast TFIID. Numbers indicate the Taf subunit name and the three lobes observed in EM are shown. Adapted from Cler, E., G. Papai, P. Schultz, and I. Davidson, 2009, Recent advances in understanding the structure and function of general transcription factor TFIID. Cell. Mol. Life Sci. 66: 2123–2134; with kind permission from Springer Science+Business Media B.V.
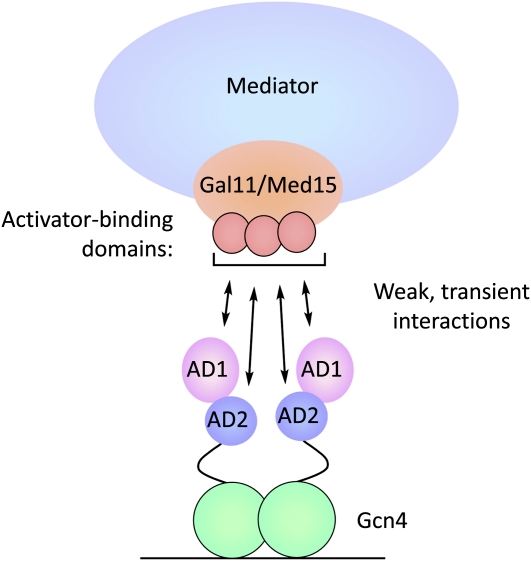
Model for Gcn4–Gal11 binding. Gcn4 contains tandem acidic activation domains and binds DNA as a dimer. Both activation domains contact at least three common activator-binding domains on Gal11/Med15, each of which contributes additively to activated transcription (Herbig et al. 2010). Activator–Gal11 binding has micromolar affinity and, for those sites measured, a half life of less than one millisecond (Jedidi et al. 2010; Brzovic et al. 2011). In this model, both Gcn4 activation domains rapidly sample the Gal11 activator-binding domains, and Mediator is recruited to the regulatory region without a stable high-affinity activator–target interaction. This binding mode can be scaled to increase Mediator recruitment by increasing the number of activator-binding sites at the promoter.
References
-
- Agricola E., Verdone L., Xella B., Di Mauro E., Caserta M., 2004. Common chromatin architecture, common chromatin remodeling, and common transcription kinetics of Adr1-dependent genes in Saccharomyces cerevisiae. Biochemistry 43: 8878–8884 - PubMed
-
- Agricola E., Verdone L., Di Mauro E., Caserta M., 2006. H4 acetylation does not replace H3 acetylation in chromatin remodelling and transcription activation of Adr1-dependent genes. Mol. Microbiol. 62: 1433–1446 - PubMed
-
- Albert I., Mavrich T. N., Tomsho L. P., Qi J., Zanton S. J., et al. , 2007. Translational and rotational settings of H2A.Z nucleosomes across the Saccharomyces cerevisiae genome. Nature 446: 572–576 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
