

Sequentially acting Sox transcription factors in neural lineage development
- PMID: 22085726
- PMCID: PMC3243056
- DOI: 10.1101/gad.176008.111
Sequentially acting Sox transcription factors in neural lineage development
Abstract
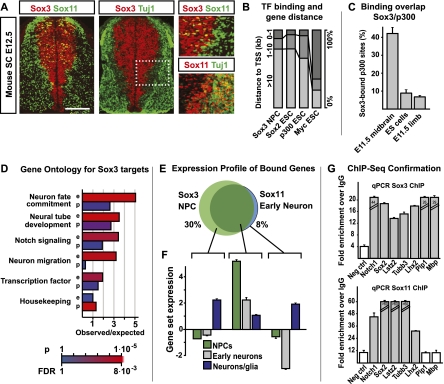
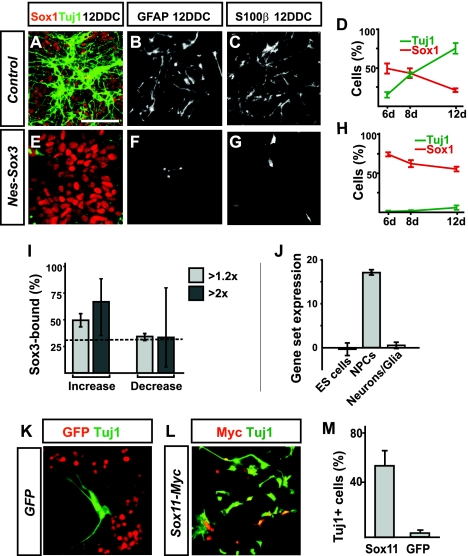
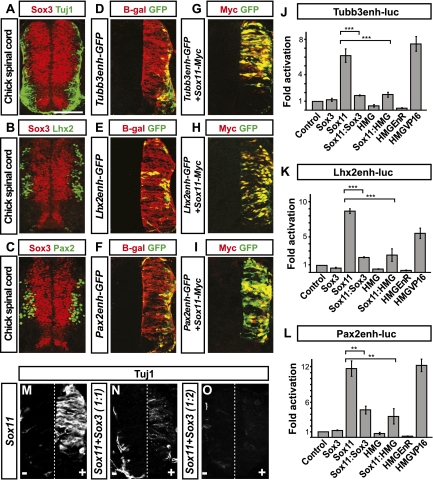
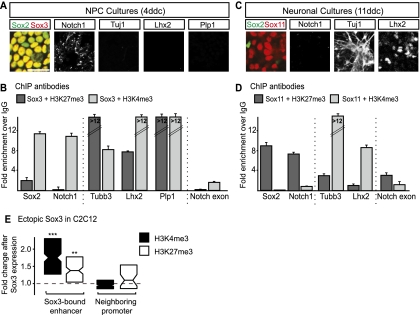
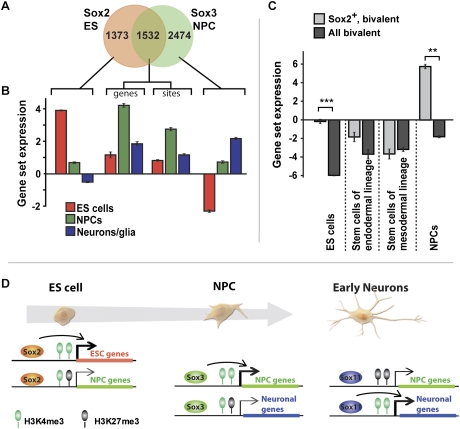
Pluripotent embryonic stem (ES) cells can generate all cell types, but how cell lineages are initially specified and maintained during development remains largely unknown. Different classes of Sox transcription factors are expressed during neurogenesis and have been assigned important roles from early lineage specification to neuronal differentiation. Here we characterize the genome-wide binding for Sox2, Sox3, and Sox11, which have vital functions in ES cells, neural precursor cells (NPCs), and maturing neurons, respectively. The data demonstrate that Sox factor binding depends on developmental stage-specific constraints and reveal a remarkable sequential binding of Sox proteins to a common set of neural genes. Interestingly, in ES cells, Sox2 preselects for neural lineage-specific genes destined to be bound and activated by Sox3 in NPCs. In NPCs, Sox3 binds genes that are later bound and activated by Sox11 in differentiating neurons. Genes prebound by Sox proteins are associated with a bivalent chromatin signature, which is resolved into a permissive monovalent state upon binding of activating Sox factors. These data indicate that a single key transcription factor family acts sequentially to coordinate neural gene expression from the early lineage specification in pluripotent cells to later stages of neuronal development.
Figures
Comment in
-
SOX after SOX: SOXession regulates neurogenesis.Genes Dev. 2011 Dec 1;25(23):2423-8. doi: 10.1101/gad.181487.111. Genes Dev. 2011. PMID: 22156204 Free PMC article.
References
-
- Altschul SF, Erickson BW 1985. Significance of nucleotide sequence alignments: A method for random sequence permutation that preserves dinucleotide and codon usage. Mol Biol Evol 2: 526–538 - PubMed
-
- Andersson E, Tryggvason U, Deng Q, Friling S, Alekseenko Z, Robert B, Perlmann T, Ericson J 2006. Identification of intrinsic determinants of midbrain dopamine neurons. Cell 124: 393–405 - PubMed
-
- Bailey TL, Elkan C 1994. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc Int Conf Intell Syst Mol Biol 2: 28–36 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous