

The effects of idebenone on mitochondrial bioenergetics
- PMID: 22086148
- PMCID: PMC3265671
- DOI: 10.1016/j.bbabio.2011.10.012
The effects of idebenone on mitochondrial bioenergetics
Abstract
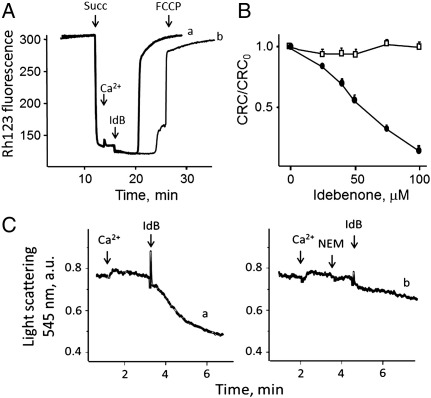
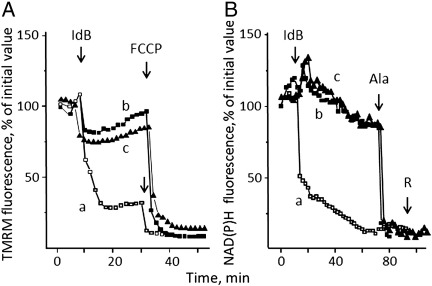
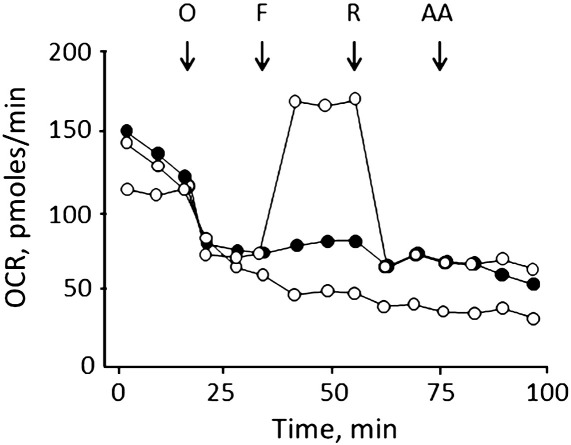
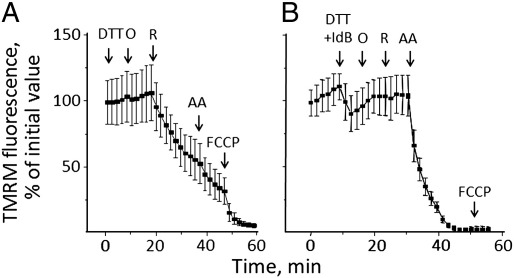
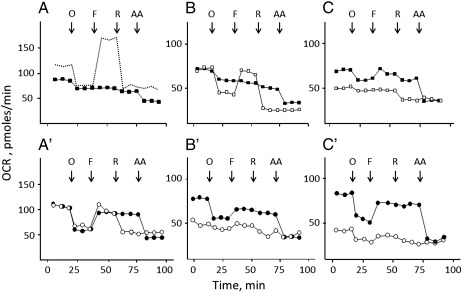
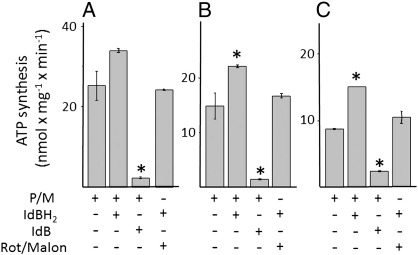
We have studied the effects of idebenone on mitochondrial function in cybrids derived from one normal donor (HQB17) and one patient harboring the G3460A/MT-ND1 mutation of Leber's Hereditary Optic Neuropathy (RJ206); and in XTC.UC1 cells bearing a premature stop codon at amino acid 101 of MT-ND1 that hampers complex I assembly. Addition of idebenone to HQB17 cells caused mitochondrial depolarization and NADH depletion, which were inhibited by cyclosporin (Cs) A and decylubiquinone, suggesting an involvement of the permeability transition pore (PTP). On the other hand, addition of dithiothreitol together with idebenone did not cause PTP opening and allowed maintenance of the mitochondrial membrane potential even in the presence of rotenone. Addition of dithiothreitol plus idebenone, or of idebenol, to HQB17, RJ206 and XTC.UC1 cells sustained membrane potential in intact cells and ATP synthesis in permeabilized cells even in the presence of rotenone and malonate, and restored a good level of coupled respiration in complex I-deficient XTC.UC1 cells. These findings demonstrate that idebenol can feed electrons at complex III. If the quinone is maintained in the reduced state, a task that in some cell types appears to be performed by dicoumarol-sensitive NAD(P)H:quinone oxidoreductase 1 [Haefeli et al. (2011) PLoS One 6, e17963], electron transfer to complex III may allow reoxidation of NADH in complex I deficiencies.
Copyright © 2011 Elsevier B.V. All rights reserved.
Figures

Similar articles
-
The idebenone metabolite QS10 restores electron transfer in complex I and coenzyme Q defects.Biochim Biophys Acta Bioenerg. 2018 Sep;1859(9):901-908. doi: 10.1016/j.bbabio.2018.04.006. Epub 2018 Apr 22. Biochim Biophys Acta Bioenerg. 2018. PMID: 29694828
-
Idebenone Has Distinct Effects on Mitochondrial Respiration in Cortical Astrocytes Compared to Cortical Neurons Due to Differential NQO1 Activity.J Neurosci. 2020 Jun 3;40(23):4609-4619. doi: 10.1523/JNEUROSCI.1632-17.2020. Epub 2020 Apr 29. J Neurosci. 2020. PMID: 32350039 Free PMC article.
-
Respiratory complex I dysfunction due to mitochondrial DNA mutations shifts the voltage threshold for opening of the permeability transition pore toward resting levels.J Biol Chem. 2009 Jan 23;284(4):2045-52. doi: 10.1074/jbc.M807321200. Epub 2008 Dec 1. J Biol Chem. 2009. PMID: 19047048
-
Idebenone and neuroprotection: antioxidant, pro-oxidant, or electron carrier?J Bioenerg Biomembr. 2015 Apr;47(1-2):111-8. doi: 10.1007/s10863-014-9571-y. Epub 2014 Sep 28. J Bioenerg Biomembr. 2015. PMID: 25262284 Free PMC article. Review.
-
Border between natural product and drug: comparison of the related benzoquinones idebenone and coenzyme Q10.Redox Biol. 2015;4:289-95. doi: 10.1016/j.redox.2015.01.009. Epub 2015 Jan 14. Redox Biol. 2015. PMID: 25625583 Free PMC article. Review.
Cited by
-
Natural Product Screening Reveals Naphthoquinone Complex I Bypass Factors.PLoS One. 2016 Sep 13;11(9):e0162686. doi: 10.1371/journal.pone.0162686. eCollection 2016. PLoS One. 2016. PMID: 27622560 Free PMC article.
-
Gamma rays induce a p53-independent mitochondrial biogenesis that is counter-regulated by HIF1α.Cell Death Dis. 2013 Jun 13;4(6):e663. doi: 10.1038/cddis.2013.187. Cell Death Dis. 2013. PMID: 23764844 Free PMC article.
-
Idebenone: A Review in Leber's Hereditary Optic Neuropathy.Drugs. 2016 May;76(7):805-13. doi: 10.1007/s40265-016-0574-3. Drugs. 2016. PMID: 27071925
-
The optic nerve: a "mito-window" on mitochondrial neurodegeneration.Mol Cell Neurosci. 2013 Jul;55(100):62-76. doi: 10.1016/j.mcn.2012.08.004. Epub 2012 Aug 15. Mol Cell Neurosci. 2013. PMID: 22960139 Free PMC article. Review.
-
Modeling Reactive Oxygen Species-Induced Axonal Loss in Leber Hereditary Optic Neuropathy.Biomolecules. 2022 Oct 2;12(10):1411. doi: 10.3390/biom12101411. Biomolecules. 2022. PMID: 36291620 Free PMC article.
References
-
- Geromel V., Darin N., Chrétien D., Bénit P., DeLonlay P., Rötig A., Munnich A., Rustin P. Coenzyme Q10 and idebenone in the therapy of respiratory chain diseases: rationale and comparative benefits. Mol. Genet. Metab. 2002;77:21–30. - PubMed
-
- Bentinger M., Tekle M., Dallner G. Coenzyme Q—biosynthesis and functions. Biochem. Biophys. Res. Commun. 2010;396:74–79. - PubMed
-
- Rotig A., Appelkvist E.L., Geromel V., Chretien D., Kadhom N., Edery P., Lebideau M., Dallner G., Munnich A., Ernster L., Rustin P. Quinone-responsive multiple respiratory-chain dysfunction due to widespread coenzyme Q10 deficiency. Lancet. 2000;356:391–395. - PubMed
-
- Ernster L., Dallner G. Biochemical, physiological and medical aspects of ubiquinone function. Biochim. Biophys. Acta. 1995;1271:195–204. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
