

Review
doi: 10.1038/nrm3226.
Tail-anchored membrane protein insertion into the endoplasmic reticulum
Affiliations
- PMID: 22086371
- PMCID: PMC3760496
- DOI: 10.1038/nrm3226
Item in Clipboard
Review
Tail-anchored membrane protein insertion into the endoplasmic reticulum
Nat Rev Mol Cell Biol.
.
Abstract
Membrane proteins are inserted into the endoplasmic reticulum (ER) by two highly conserved parallel pathways. The well-studied co-translational pathway uses signal recognition particle (SRP) and its receptor for targeting and the SEC61 translocon for membrane integration. A recently discovered post-translational pathway uses an entirely different set of factors involving transmembrane domain (TMD)-selective cytosolic chaperones and an accompanying receptor at the ER. Elucidation of the structural and mechanistic basis of this post-translational membrane protein insertion pathway highlights general principles shared between the two pathways and key distinctions unique to each.
Figures
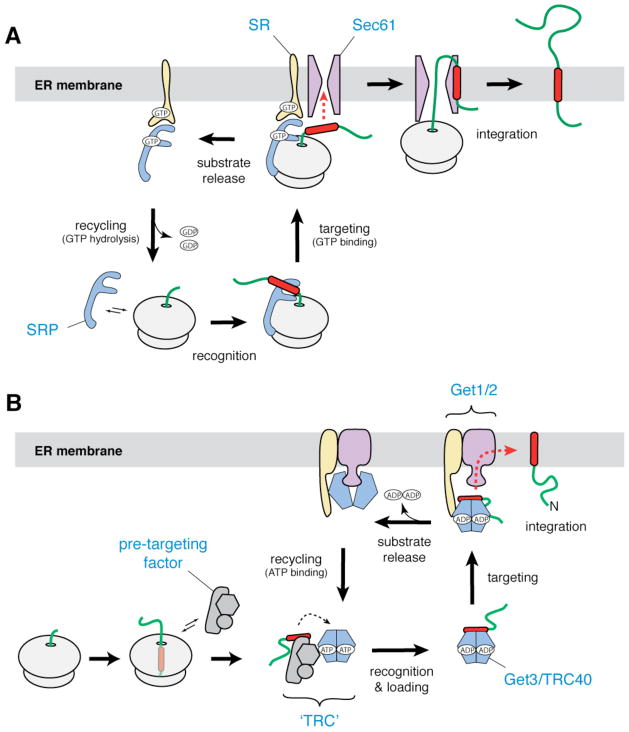
a | In the co-translational pathway for insertion of ER membrane proteins, SRP (blue) recognizes the hydrophobic signal peptide (red) of the nascent chain as it emerges from a translating ribosome. The ribosome–nascent chain–SRP complex is targeted to the membrane by a GTP-dependent interaction with the SRP receptor (yellow), resulting in release of the signal peptide and docking of the ribosome–nascent chain complex to the Sec61 translocon (magenta). Translation then resumes, and the nascent polypeptide is inserted into the membrane bilayer. After GTP hydrolysis, SRP is recycled to the cytosol. b | In the post-translational pathway for insertion of tail-anchored (TA) ER membrane proteins, a soluble pre-targeting complex (grey) captures the hydrophobic TMD (red) of the TA substrate after it emerges from the ribosomal exit channel. After loading onto Get3/TRC40 (shown as a blue–green dimer), the TA substrate is delivered to the ER membrane by interaction with the Get1–Get2 receptor complex (yellow and magenta). Upon ATP hydrolysis, the TA substrate is released for insertion into the bilayer. ATP binding recycles Get3/TRC40 back to the cytosol.
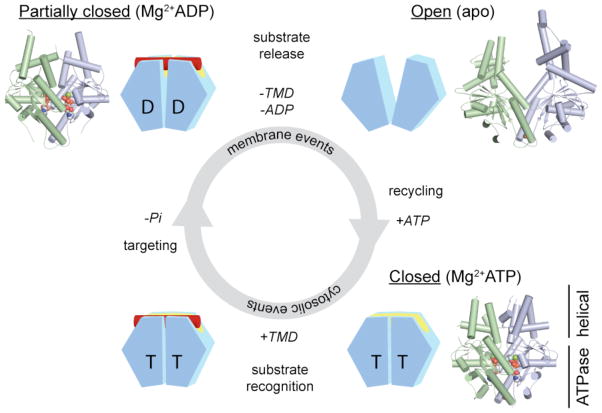
Each Get3 monomer comprises two distinct regions: an ATPase- and an α-helical subdomain. In the presence of ATP, the Get3 helical subdomains become intimately associated, forming an extended composite hydrophobic groove (see FIG. 3) that recognizes and binds to the TMD of a TA substrate (red). ATP hydrolysis, which occurs at some stage prior to release of the TA substrate at the ER membrane, produces an ADP-bound Get3 homodimer that is partially closed. Following release of the TA substrate and ADP, Get3 shifts back to an open conformation. Subsequently, ATP binding allows recycling of Get3 back to the cytosol in a closed conformation. The insets show crystal structures of the fungal Get3 homodimer in the nucleotide-free (PDB ID 2WOO), Mg2+ADP-bound (PDB ID 3IQX) and Mg2+ADP•AlF4−-bound (PDB ID 2WOJ) states.
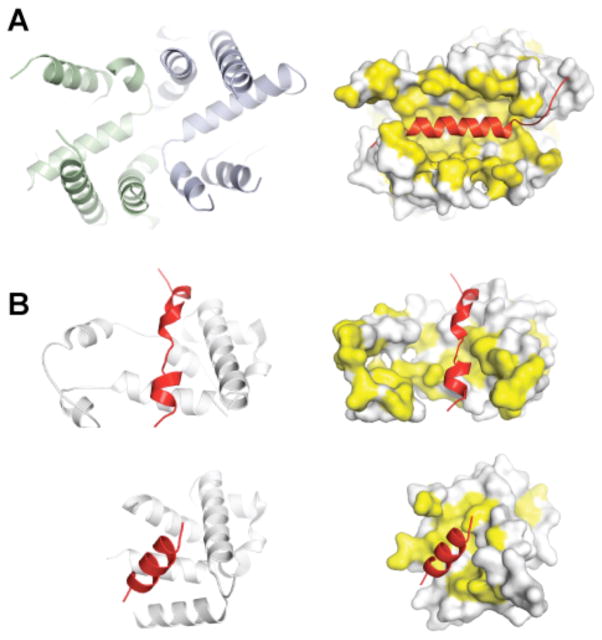
a | A large groove in Get3 is formed by association of the two α-helical subdomains (left panel, green and blue) in the ATP-bound, closed dimer conformation. This composite groove presents a large hydrophobic surface for binding to the TMD of a TA protein substrate. The TA protein Sec61β (shown on the right side in red; PDB ID 1RHZ) is modelled into the groove; hydrophobic surfaces are in yellow. b | Crystal structures of the methionine-rich M-domain of SRP54 bound to a signal peptide (shown in red) (top, PDB ID 3KL4; bottom, PDB ID 3NBD). Note the different, but overlapping peptide binding sites. For both the post-translational TA protein pathway (a) and the co-translational pathway (b), the dynamic properties of these helical, hydrophobic scaffolds probably allows them to accommodate different sequences during targeting.

The TMD recognition complex (TRC) consists of a stable core complex (depicted in grey) and several dynamically associated components. In budding yeast, the TRC core consists of Get4 and Get5, with Get4 recruiting Get3, and Get5 recruiting Sgt2. Sgt2 can additionally recruit other chaperones. In mammals, TRC35 (which is homologous to Get4) and Ubl4A (which is homologous to Get5) are in a complex with Bag6, which probably recruits SGTA (which is homologous to Sgt2). Engagement of the TRC by a substrate (probably bound to Sgt2/SGTA) results in its sorting among any of several potential TMD-binding proteins (dashed arrows). This sorting is presumably dictated by a combination of substrate features and availability of the binding partners. The substrate can therefore emerge from the TRC bound to any of multiple binding partners, each of which imparts a specific downstream fate. Get3/TRC40 association mediates ER targeting, whereas Bag6 binding can recruit an E3 ubiquitin ligase that mediates substrate degradation. The fate of other complexes are not understood but could include targeting to other destinations including the mitochondria.
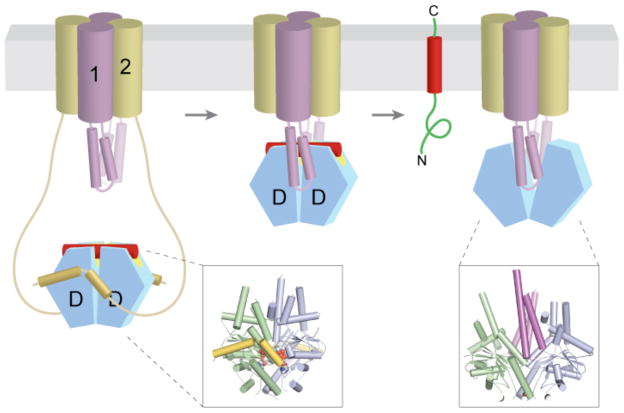
Nucleotide (either ADP or ATP) and TA substrate (red)-bound Get3 (blue and green) is captured at the ER membrane by the long, flexible amino termini of Get2 (yellow). After ATP hydrolysis, the Get3–TA substrate complex is transferred to Get1, which wedges open the composite hydrophobic groove to promote TA substrate (and ADP) release. Finally, ATP re-binding dissociates the stable Get1–Get3 post-insertion complex to recycle Get3 back to the cytosol. Although depicted here as a stable heterotetramer, the stoichiometry and subunit composition of the Get1–Get2 receptor complex is not known. The insets show crystal structures of Mg2+ADP•AlF4−-bound Get3 in complex with the cytosolic fragment of Get2 (left, PDB ID 3ZS9), and of nucleotide-free Get3 in complex with the cytosolic fragment of Get1 (right, PDB ID 3ZS8).
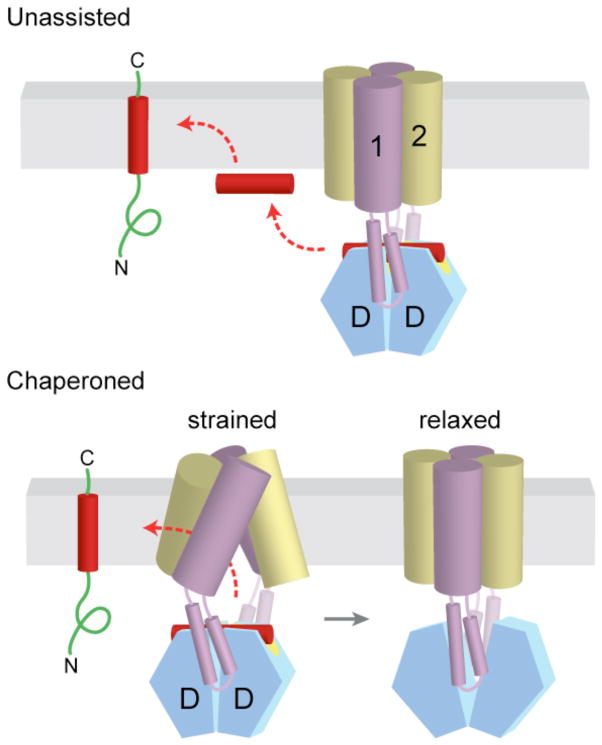
a | After release from Get3, the TA substrate might transiently associate with the membrane surface before inserting spontaneously into the lipid bilayer. b | Alternatively, the transmembrane domains (TMDs) of Get1 and Get2 may interact directly with the TA substrate to ‘chaperone’ it into the bilayer. In the model shown here, binding of two Get1 subunits to the partially closed targeting complex results in a ‘strained’ configuration of the Get1–Get2 receptor complex. This arrangement might provide a hydrophobic gate through which the TMD could diffuse into the bilayer. ‘Wedging open’ the Get3 dimer for substrate release would then allow the Get1–Get2 receptor complex to ‘relax’ back into a low-energy configuration that is sealed off from the cytosol. Thus, conformational changes in Get3 might be coupled to those of the receptor TMDs to promote TA substrate integration.
References
-
- Fagerberg L, Jonasson K, von Heijne G, Uhlen M, Berglund L. Prediction of the human membrane proteome. Proteomics. 2010;10:1141–1149. - PubMed
-
- White SH, von Heijne G. Transmembrane helices before, during, and after insertion. Curr Opin Struct Biol. 2005;15:378–386. - PubMed
-
- White SH, von Heijne G. How translocons select transmembrane helices. Annu Rev Biophys. 2008;37:23–42. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
