

Nutrient sensing kinases PKA and Sch9 phosphorylate the catalytic domain of the ubiquitin-conjugating enzyme Cdc34
- PMID: 22087249
- PMCID: PMC3210133
- DOI: 10.1371/journal.pone.0027099
Nutrient sensing kinases PKA and Sch9 phosphorylate the catalytic domain of the ubiquitin-conjugating enzyme Cdc34
Abstract
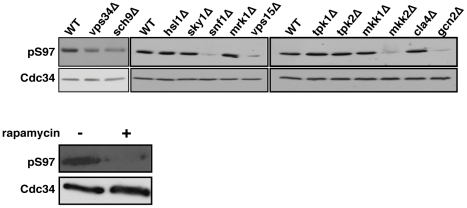
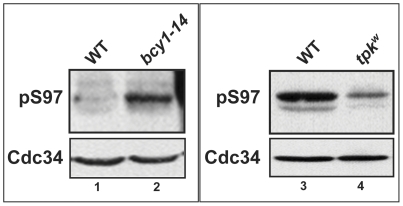
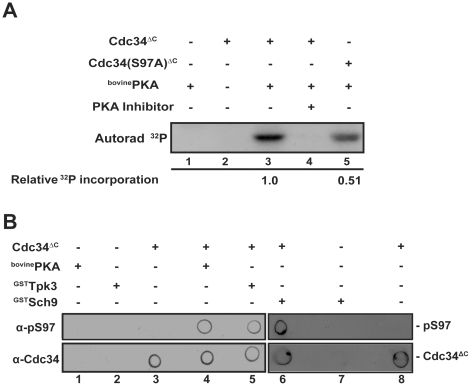
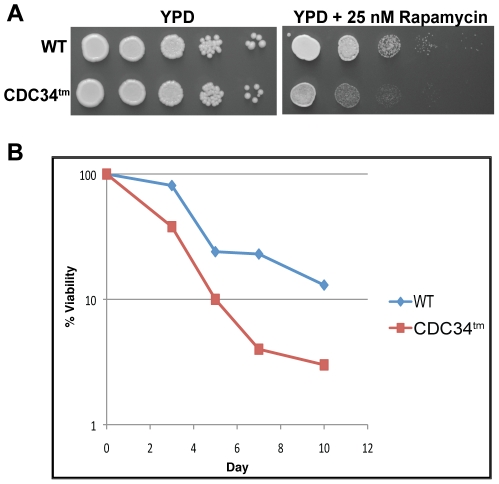
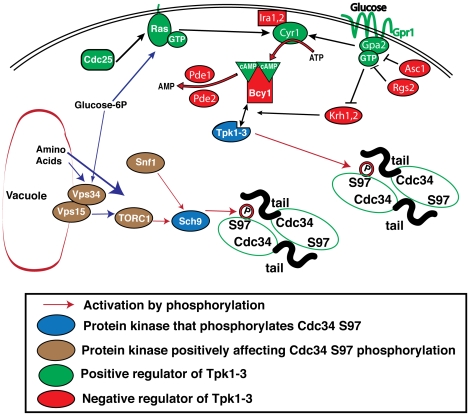
Cell division is controlled in part by the timely activation of the CDK, Cdc28, through its association with G1 and G2 cyclins. Cdc28 complexes are regulated in turn by the ubiquitin conjugating enzyme Cdc34 and SCF ubiquitin ligase complexes of the ubiquitin-proteasome system (UPS) to control the initiation of DNA replication. Here we demonstrate that the nutrient sensing kinases PKA and Sch9 phosphorylate S97 of Cdc34. S97 is conserved across species and restricted to the catalytic domain of Cdc34/Ubc7-like E2s. Cdc34-S97 phosphorylation is cell cycle regulated, elevated during active cell growth and division and decreased during cell cycle arrest. Cell growth and cell division are orchestrated to maintain cell size homeostasis over a wide range of nutrient conditions. Cells monitor changes in their environment through nutrient sensing protein kinases. Thus Cdc34 phosphorylation by PKA and Sch9 provides a direct tether between G1 cell division events and cell growth.
Conflict of interest statement
Figures

Similar articles
-
Cdc34 C-terminal tail phosphorylation regulates Skp1/cullin/F-box (SCF)-mediated ubiquitination and cell cycle progression.Biochem J. 2007 Aug 1;405(3):569-81. doi: 10.1042/BJ20061812. Biochem J. 2007. PMID: 17461777 Free PMC article.
-
Association of the disordered C-terminus of CDC34 with a catalytically bound ubiquitin.J Mol Biol. 2011 Apr 1;407(3):425-38. doi: 10.1016/j.jmb.2011.01.047. Epub 2011 Feb 4. J Mol Biol. 2011. PMID: 21296085
-
Multimodal mechanism of action for the Cdc34 acidic loop: a case study for why ubiquitin-conjugating enzymes have loops and tails.J Biol Chem. 2013 Nov 29;288(48):34882-96. doi: 10.1074/jbc.M113.509190. Epub 2013 Oct 15. J Biol Chem. 2013. PMID: 24129577 Free PMC article.
-
Cell cycle regulation by the ubiquitin pathway.FASEB J. 1997 Nov;11(13):1067-75. doi: 10.1096/fasebj.11.13.9367342. FASEB J. 1997. PMID: 9367342 Review.
-
How proteolysis drives the cell cycle.Science. 1996 Dec 6;274(5293):1652-9. doi: 10.1126/science.274.5293.1652. Science. 1996. PMID: 8939846 Review.
Cited by
-
Activating the Anaphase Promoting Complex to Enhance Genomic Stability and Prolong Lifespan.Int J Mol Sci. 2018 Jun 27;19(7):1888. doi: 10.3390/ijms19071888. Int J Mol Sci. 2018. PMID: 29954095 Free PMC article. Review.
-
The yeast protein kinase Sch9 adjusts V-ATPase assembly/disassembly to control pH homeostasis and longevity in response to glucose availability.PLoS Genet. 2017 Jun 12;13(6):e1006835. doi: 10.1371/journal.pgen.1006835. eCollection 2017 Jun. PLoS Genet. 2017. PMID: 28604780 Free PMC article.
-
Control of APC/C-dependent ubiquitin chain elongation by reversible phosphorylation.Proc Natl Acad Sci U S A. 2016 Feb 9;113(6):1540-5. doi: 10.1073/pnas.1522423113. Epub 2016 Jan 25. Proc Natl Acad Sci U S A. 2016. PMID: 26811472 Free PMC article.
-
Using the AKAR3-EV biosensor to assess Sch9p- and PKA-signalling in budding yeast.FEMS Yeast Res. 2023 Jan 4;23:foad029. doi: 10.1093/femsyr/foad029. FEMS Yeast Res. 2023. PMID: 37173282 Free PMC article.
-
DNA replication stress-induced loss of reproductive capacity in S. cerevisiae and its inhibition by caloric restriction.Cell Cycle. 2013 Apr 15;12(8):1189-200. doi: 10.4161/cc.24232. Epub 2013 Mar 21. Cell Cycle. 2013. PMID: 23518504 Free PMC article.
References
-
- Glickman MH, Ciechanover A. The ubiquitin-proteasome proteolytic pathway: destruction for the sake of construction. Physiol Rev. 2002;82:373–428. - PubMed
-
- Goebl MG, Yochem J, Jentsch S, McGrath JP, Varshavsky A, et al. The yeast cell cycle gene CDC34 encodes a ubiquitin-conjugating enzyme. Science. 1988;241:1331–1335. - PubMed
-
- Mathias N, Steussy CN, Goebl MG. An essential domain within Cdc34p is required for binding to a complex containing Cdc4p and Cdc53p in Saccharomyces cerevisiae. J Biol Chem. 1998;273:4040–4045. - PubMed
-
- Deshaies RJ. SCF and Cullin/Ring H2-based ubiquitin ligases. Annu Rev Cell Dev Biol. 1999;15:435–467. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
