

Intermediates in the transformation of phosphonates to phosphate by bacteria
- PMID: 22089136
- PMCID: PMC3245791
- DOI: 10.1038/nature10622
Intermediates in the transformation of phosphonates to phosphate by bacteria
Abstract
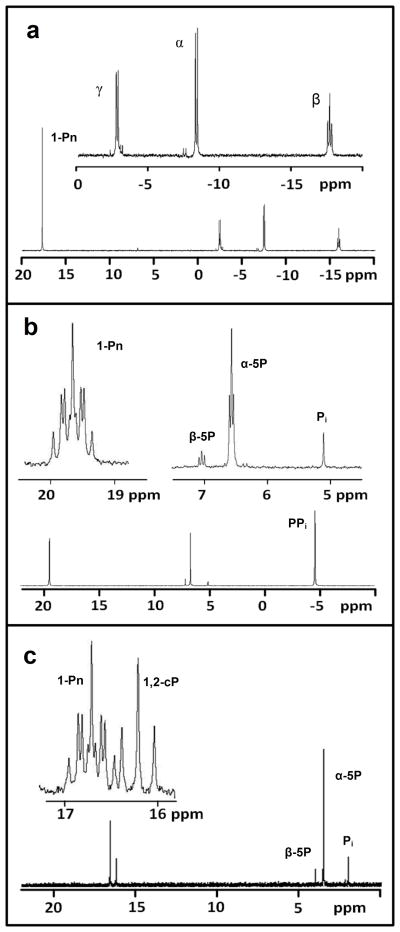
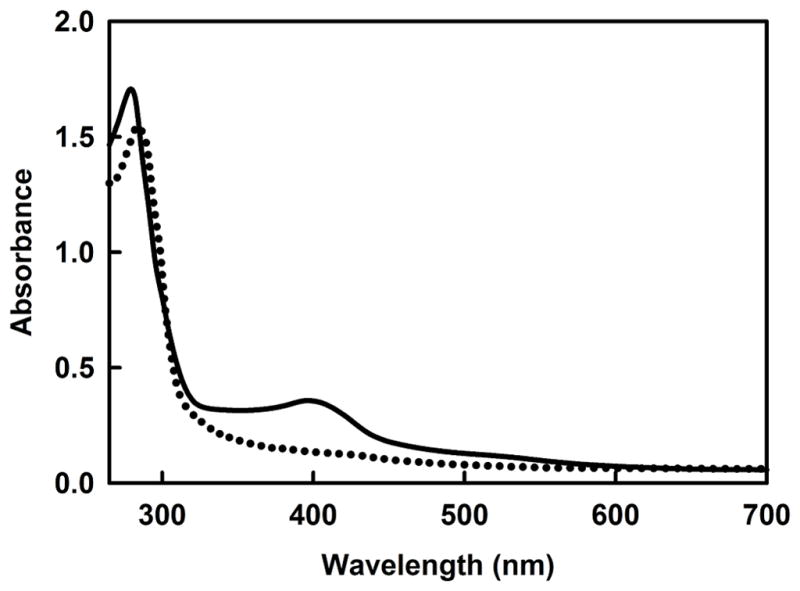
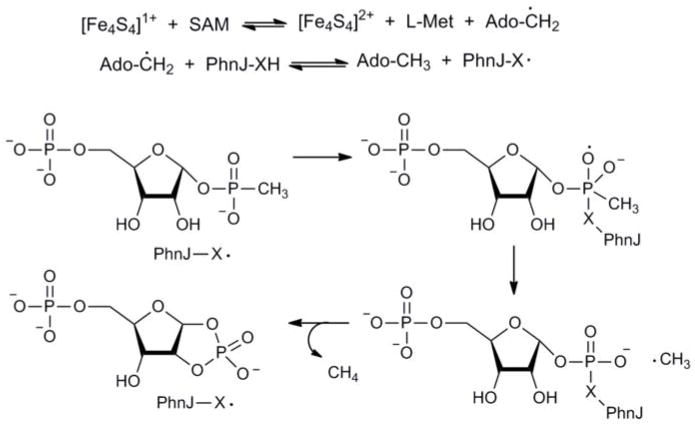
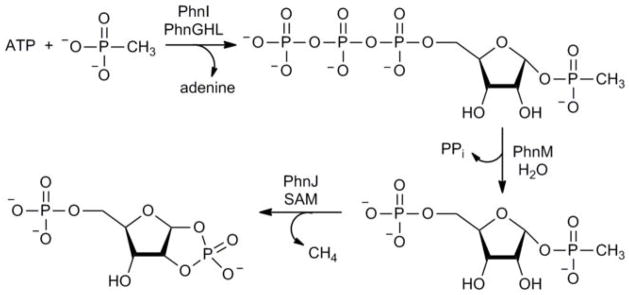
Phosphorus is an essential element for all known forms of life. In living systems, phosphorus is an integral component of nucleic acids, carbohydrates and phospholipids, where it is incorporated as a derivative of phosphate. However, most Gram-negative bacteria have the capability to use phosphonates as a nutritional source of phosphorus under conditions of phosphate starvation. In these organisms, methylphosphonate is converted to phosphate and methane. In a formal sense, this transformation is a hydrolytic cleavage of a carbon-phosphorus (C-P) bond, but a general enzymatic mechanism for the activation and conversion of alkylphosphonates to phosphate and an alkane has not been elucidated despite much effort for more than two decades. The actual mechanism for C-P bond cleavage is likely to be a radical-based transformation. In Escherichia coli, the catalytic machinery for the C-P lyase reaction has been localized to the phn gene cluster. This operon consists of the 14 genes phnC, phnD, …, phnP. Genetic and biochemical experiments have demonstrated that the genes phnG, phnH, …, phnM encode proteins that are essential for the conversion of phosphonates to phosphate and that the proteins encoded by the other genes in the operon have auxiliary functions. There are no functional annotations for any of the seven proteins considered essential for C-P bond cleavage. Here we show that methylphosphonate reacts with MgATP to form α-D-ribose-1-methylphosphonate-5-triphosphate (RPnTP) and adenine. The triphosphate moiety of RPnTP is hydrolysed to pyrophosphate and α-D-ribose-1-methylphosphonate-5-phosphate (PRPn). The C-P bond of PRPn is subsequently cleaved in a radical-based reaction producing α-D-ribose-1,2-cyclic-phosphate-5-phosphate and methane in the presence of S-adenosyl-L-methionine. Substantial quantities of phosphonates are produced worldwide for industrial processes, detergents, herbicides and pharmaceuticals. Our elucidation of the chemical steps for the biodegradation of alkylphosphonates shows how these compounds can be metabolized and recycled to phosphate.
Conflict of interest statement
Figures
References
-
- Metcalf WW, Wanner BL. Evidence for a fourteen-gene, phnC to phnP locus for phosphonate metabolism in Escherichia coli. Gene. 1993;129:27–32. - PubMed
-
- Ahn Y, Ye Q, Cho H, Walsh CT, Floss HG. Stereochemistry of carbon-phosphorus cleavage in ethylphosphonate catalyzed by C-P lyase from Escherichia coli. J Am Chem Soc. 1992;114:7953–7954.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
