

A male sterility-associated cytotoxic protein ORF288 in Brassica juncea causes aborted pollen development
- PMID: 22090439
- PMCID: PMC3276091
- DOI: 10.1093/jxb/err355
A male sterility-associated cytotoxic protein ORF288 in Brassica juncea causes aborted pollen development
Abstract
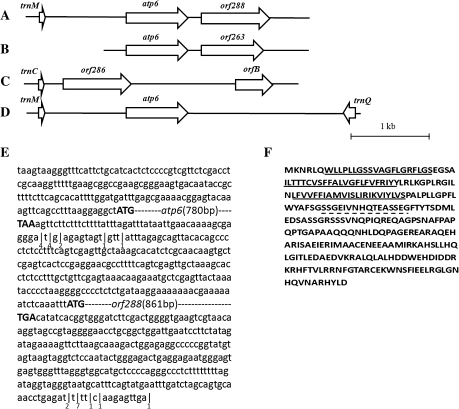
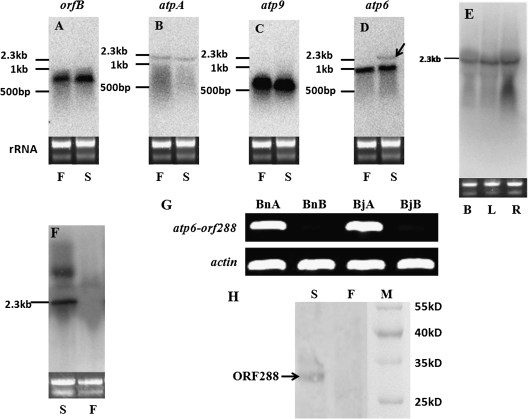
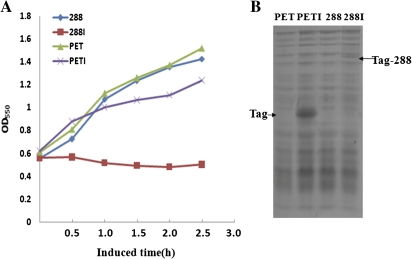
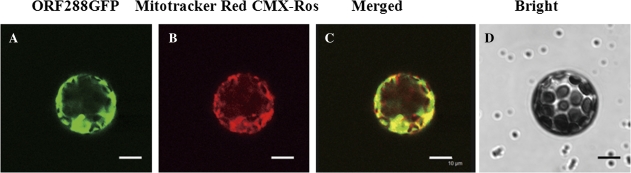
Cytoplasmic male sterility (CMS) is a widespread phenomenon in higher plants, and several studies have established that this maternally inherited defect is often associated with a mitochondrial mutant. Approximately 10 chimeric genes have been identified as being associated with corresponding CMS systems in the family Brassicaceae, but there is little direct evidence that these genes cause male sterility. In this study, a novel chimeric gene (named orf288) was found to be located downstream of the atp6 gene and co-transcribed with this gene in the hau CMS sterile line. Western blotting analysis showed that this predicted open reading frame (ORF) was translated in the mitochondria of male-sterile plants. Furthermore, the growth of Escherichia coli was significantly repressed in the presence of ORF288, which indicated that this protein is toxic to the E. coli host cells. To confirm further the function of orf288 in male sterility, the gene was fused to a mitochondrial-targeting pre-sequence under the control of the Arabidopsis APETALA3 promoter and introduced into Arabidopsis thaliana. Almost 80% of transgenic plants with orf288 failed to develop anthers. It was also found that the independent expression of orf288 caused male sterility in transgenic plants, even without the transit pre-sequence. Furthermore, transient expression of orf288 and green fluorescent protein (GFP) as a fused protein in A. thaliana protoplasts showed that ORF288 was able to anchor to mitochondria even without the external mitochondrial-targeting peptide. These observations provide important evidence that orf288 is responsible for the male sterility of hau CMS in Brassica juncea.
Figures

References
-
- Akagi H, Nakamura A, Yokozeki-Misono Y, Inagaki A, Takahashi H, Mori K, Fujimura T. Positional cloning of the rice Rf-1 gene, a restorer of BT-type cytoplasmic male sterility that encodes a mitochondria-targeting PPR protein. Theoretical and Applied Genetics. 2004;108:1449–1457. - PubMed
-
- Ashutosh Kumar P, Dinesh Kumar V, Sharma PC, Prakash S, Bhat SR. A novel orf108 co-transcribed with the atpA gene is associated with cytoplasmic male sterility in Brassica juncea carrying Moricandia arvensis cytoplasm. Plant and Cell Physiology. 2008;49:284–289. - PubMed
-
- Bellaoui M, Grelon M, Pelletier G, Budar F. The restorer Rfo gene acts post-translationally on the stability of the ORF138 Ogura CMS-associated protein in reproductive tissues of rapeseed cybrids. Plant Molecular Biology. 1999;40:893–902. - PubMed
-
- Bendtsen JD, Nielsen H, von Heijne G, Brunak S. Improved prediction of signal peptides: SignalP 3.0. Journal of Molecular Biology. 2004;340:783–795. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
