

Genome-environment interactions that modulate aging: powerful targets for drug discovery
- PMID: 22090473
- PMCID: PMC3250080
- DOI: 10.1124/pr.110.004499
Genome-environment interactions that modulate aging: powerful targets for drug discovery
Abstract
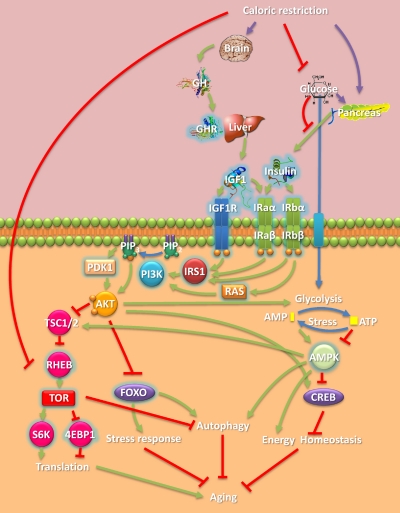
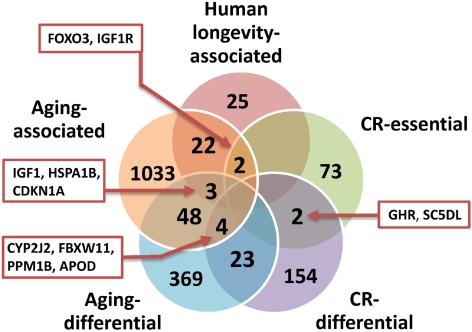
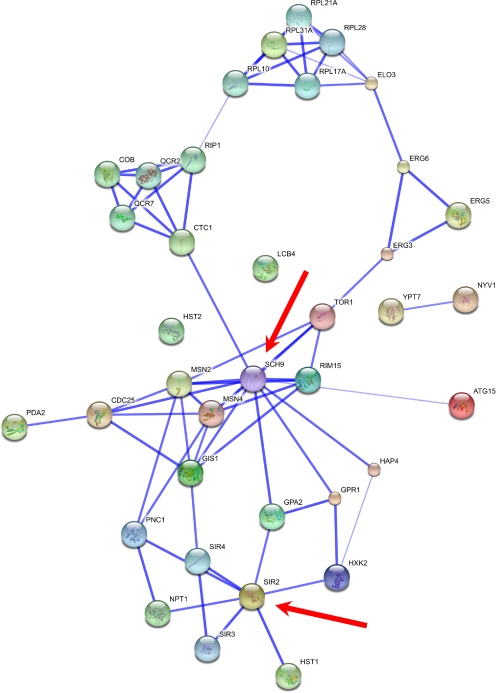
Aging is the major biomedical challenge of this century. The percentage of elderly people, and consequently the incidence of age-related diseases such as heart disease, cancer, and neurodegenerative diseases, is projected to increase considerably in the coming decades. Findings from model organisms have revealed that aging is a surprisingly plastic process that can be manipulated by both genetic and environmental factors. Here we review a broad range of findings in model organisms, from environmental to genetic manipulations of aging, with a focus on those with underlying gene-environment interactions with potential for drug discovery and development. One well-studied dietary manipulation of aging is caloric restriction, which consists of restricting the food intake of organisms without triggering malnutrition and has been shown to retard aging in model organisms. Caloric restriction is already being used as a paradigm for developing compounds that mimic its life-extension effects and might therefore have therapeutic value. The potential for further advances in this field is immense; hundreds of genes in several pathways have recently emerged as regulators of aging and caloric restriction in model organisms. Some of these genes, such as IGF1R and FOXO3, have also been associated with human longevity in genetic association studies. The parallel emergence of network approaches offers prospects to develop multitarget drugs and combinatorial therapies. Understanding how the environment modulates aging-related genes may lead to human applications and disease therapies through diet, lifestyle, or pharmacological interventions. Unlocking the capacity to manipulate human aging would result in unprecedented health benefits.
Figures
References
-
- Anisimov VN, Berstein LM, Egormin PA, Piskunova TS, Popovich IG, Zabezhinski MA, Tyndyk ML, Yurova MV, Kovalenko IG, Poroshina TE, et al. (2008) Metformin slows down aging and extends life span of female SHR mice. Cell Cycle 7:2769–2773 - PubMed
-
- Ayyadevara S, Alla R, Thaden JJ, Shmookler Reis RJ. (2008) Remarkable longevity and stress resistance of nematode PI3K-null mutants. Aging Cell 7:13–22 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
