

Analgesic conotoxins: block and G protein-coupled receptor modulation of N-type (Ca(V) 2.2) calcium channels
- PMID: 22091786
- PMCID: PMC3417482
- DOI: 10.1111/j.1476-5381.2011.01781.x
Analgesic conotoxins: block and G protein-coupled receptor modulation of N-type (Ca(V) 2.2) calcium channels
Abstract
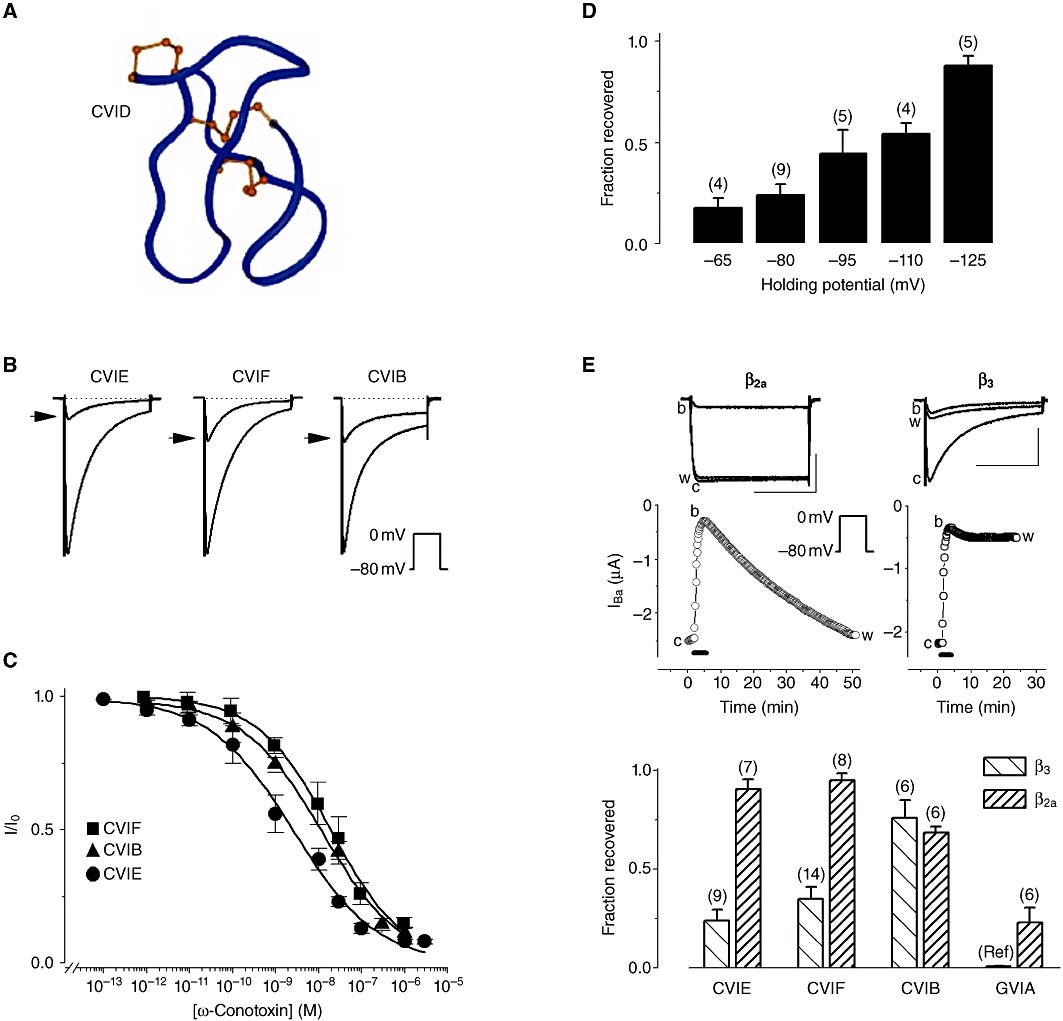
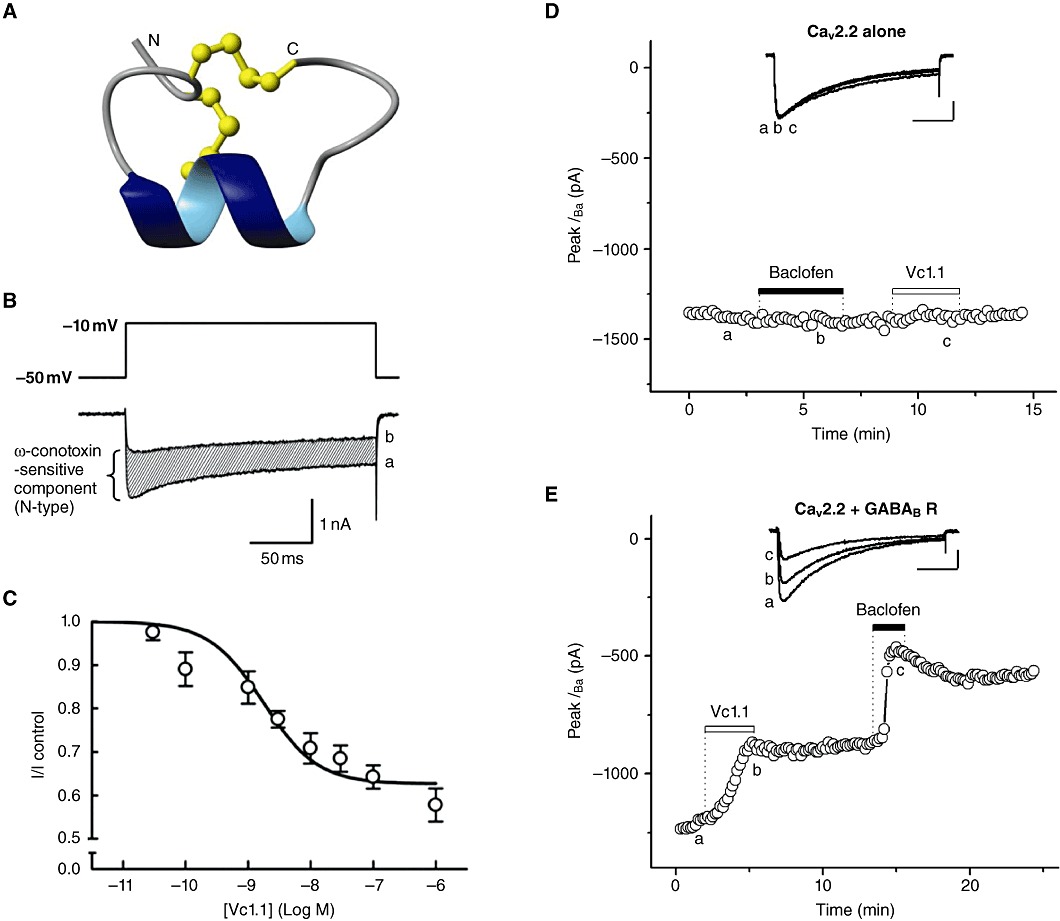
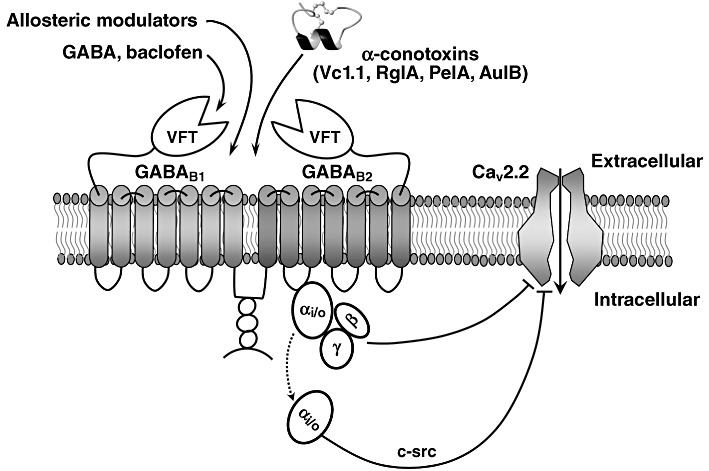
Conotoxins (conopeptides) are small disulfide bonded peptides from the venom of marine cone snails. These peptides target a wide variety of membrane receptors, ion channels and transporters, and have enormous potential for a range of pharmaceutical applications. Structurally related ω-conotoxins bind directly to and selectively inhibit neuronal (N)-type voltage-gated calcium channels (VGCCs) of nociceptive primary afferent neurones. Among these, ω-conotoxin MVIIA (Prialt) is approved by the Food and Drug Administration (FDA) as an alternative intrathecal analgesic for the management of chronic intractable pain, particularly in patients refractory to opioids. A series of newly discovered ω-conotoxins from Conus catus, including CVID-F, are potent and selective antagonists of N-type VGCCs. In spinal cord slices, these peptides reversibly inhibit excitatory synaptic transmission between primary afferents and dorsal horn superficial lamina neurones, and in the rat partial sciatic nerve ligation model of neuropathic pain, significantly reduce allodynic behaviour. Another family of conotoxins, the α-conotoxins, are competitive antagonists of mammalian nicotinic acetylcholine receptors (nAChRs). α-Conotoxins Vc1.1 and RgIA possess two disulfide bonds and are currently in development as a treatment for neuropathic pain. It was initially proposed that the primary target of these peptides is the α9α10 neuronal nAChR. Surprisingly, however, α-conotoxins Vc1.1, RgIA and PeIA more potently inhibit N-type VGCC currents via a GABA(B) GPCR mechanism in rat sensory neurones. This inhibition is largely voltage-independent and involves complex intracellular signalling. Understanding the molecular mechanisms of conotoxin action will lead to new ways to regulate VGCC block and modulation in normal and diseased states of the nervous system.
© 2011 The Authors. British Journal of Pharmacology © 2011 The British Pharmacological Society.
Figures
References
-
- Abbott JR, Litzinger MJ. Different ω-conotoxins mark the development of Swiss Webster mouse cortex suggesting N-type voltage sensitive calcium channel subtypes. Int J Dev Neurosci. 1994;12:43–47. - PubMed
-
- Adams DJ, Alewood PF, Craik DJ, Drinkwater RD, Lewis RJ. Conotoxins and their potential pharmaceutical applications. Drug Dev Res. 1999;46:219–234.
-
- Adams DJ, Smith AB, Schroeder CI, Yasuda T, Lewis RJ. ω-Conotoxin CVID inhibits a pharmacologically distinct voltage-sensitive calcium channel associated with transmitter release from preganglionic nerve terminals. J Biol Chem. 2003;278:4057–4062. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
