

Review
doi: 10.1111/j.1574-6976.2011.00310.x.
Epub 2011 Oct 25.
Recent progress in Bacillus subtilis sporulation
Affiliations
- PMID: 22091839
- PMCID: PMC3237856
- DOI: 10.1111/j.1574-6976.2011.00310.x
Item in Clipboard
Review
Recent progress in Bacillus subtilis sporulation
FEMS Microbiol Rev.
2012 Jan.
Abstract
The Gram-positive bacterium Bacillus subtilis can initiate the process of sporulation under conditions of nutrient limitation. Here, we review some of the last 5 years of work in this area, with a particular focus on the decision to initiate sporulation, DNA translocation, cell-cell communication, protein localization and spore morphogenesis. The progress we describe has implications not only just for the study of sporulation but also for other biological systems where homologs of sporulation-specific proteins are involved in vegetative growth.
© 2011 Federation of European Microbiological Societies. Published by Blackwell Publishing Ltd. All rights reserved.
Figures
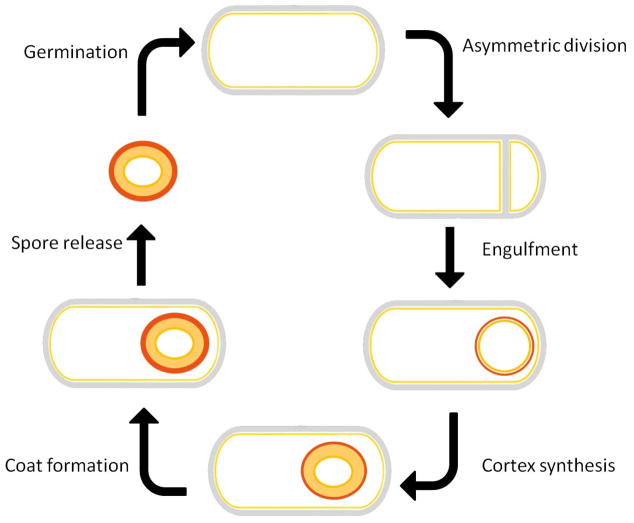
Upon nutrient limitation, B. subtilis ceases growth and initiates sporulation. An asymmetric cell division generates a smaller cell and a larger cell. Following completion of DNA segregation, the larger cell (the mother cell) proceeds to engulf the smaller cell (the forespore). During the initiation of spore metabolic dormancy and compaction of the spore DNA, the mother cell mediates the development of the forespore into the spore through the production of the spore cortex and the inner and outer coat. Upon completion of this process, the mother cell lyses, releasing the mature spore. When the dormant spore encounters an appropriate environmental stimulus, it initiates the process of germination that can result in the re-initiation of vegetative growth if sufficient nutrients are present.
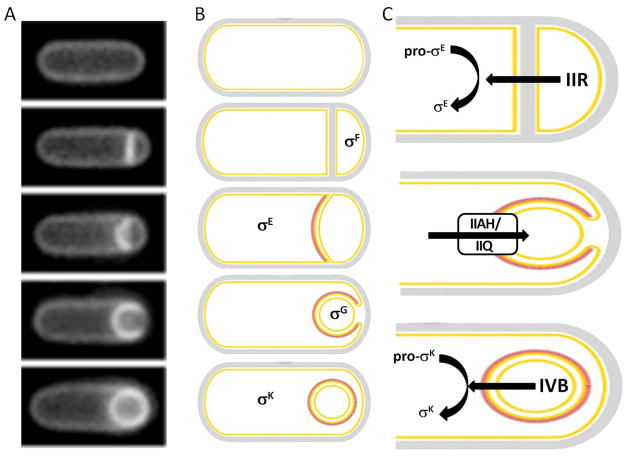
(A) Morphological asymmetry. Following entry into sporulation, the cells form an asymmetric septum, which then proceeds to curve out; it eventually becomes entirely distorted and surrounds the smaller cell. This process of engulfment is completed by the loss of attachment between the forespore and the mother cell. (B) Genetic asymmetry. These morphological changes are accompanied by activation of specific transcription factors in each compartment. Initially, σF is activated only in the forespore, followed by σE in the mother cell, σG in the forespore, and finally σK in the mother cell. (C) Intercellular signaling cascade underlying genetic asymmetry. (i) The SpoIIR protein that is under control of σF is produced and secreted by the forespore and activates proteolytic processing of pro-σE→σE in the mother cell. (ii) An as yet unknown signal, produced in the mother cell under control of σE, enters the forespore presumably via the SpoIIIAH/SpoIIQ pore and activates σG. (iii) The SpoIVB protein produced under the control of σG in the forespore activates pro-σK→σK proteolytic processing in the mother cell via an interaction with the SpoIVFA/SpoIVFB/BofA complex
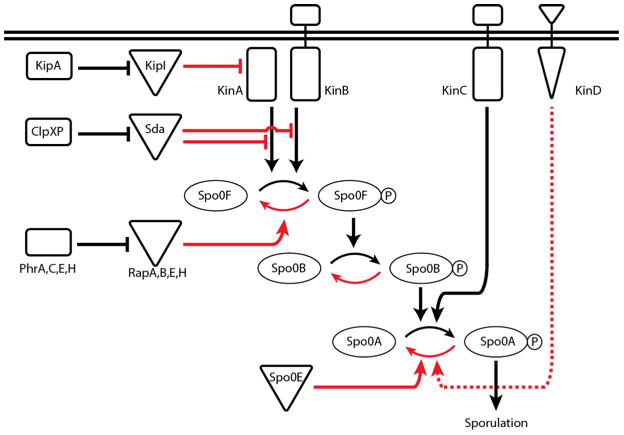
SpoOA phosphorylation is controlled by a phosphorelay comprised of several kinases and numerous intermediaries. Interactions that ultimately work against SpoOA phosphorylation appear in red while interactions that lead towards SpoOA~P are presented in black. At the beginning of the phosphorelay, the KinA and KinB kinases respond to as yet unknown signals by autophosphorylating and subsequently transferring phosphate to SpoOF. SpoOF phosphorylates SpoOB, which passes phosphate on to SpoOA. A third kinase, KinC, bypasses the phosphorelay (i.e. SpoOF and SpoOB) and acts directly on SpoOA. A number of phosphatases (RapA,B,E,&H and SpoOE) and kinase inhibitors (Sda and KipI) act at various steps in the pathway to directly or indirectly antagonize SpoOA phosphorylation. These inhibitors are subject to their own regulation. KinD has been shown to work against SpoOA phosphorylation though it has not yet been determined at which step this regulation occurs (it is therefore graphed as a dashed line). This entire network of interactions is ultimately represented by the phosphorylation state of SpoOA.
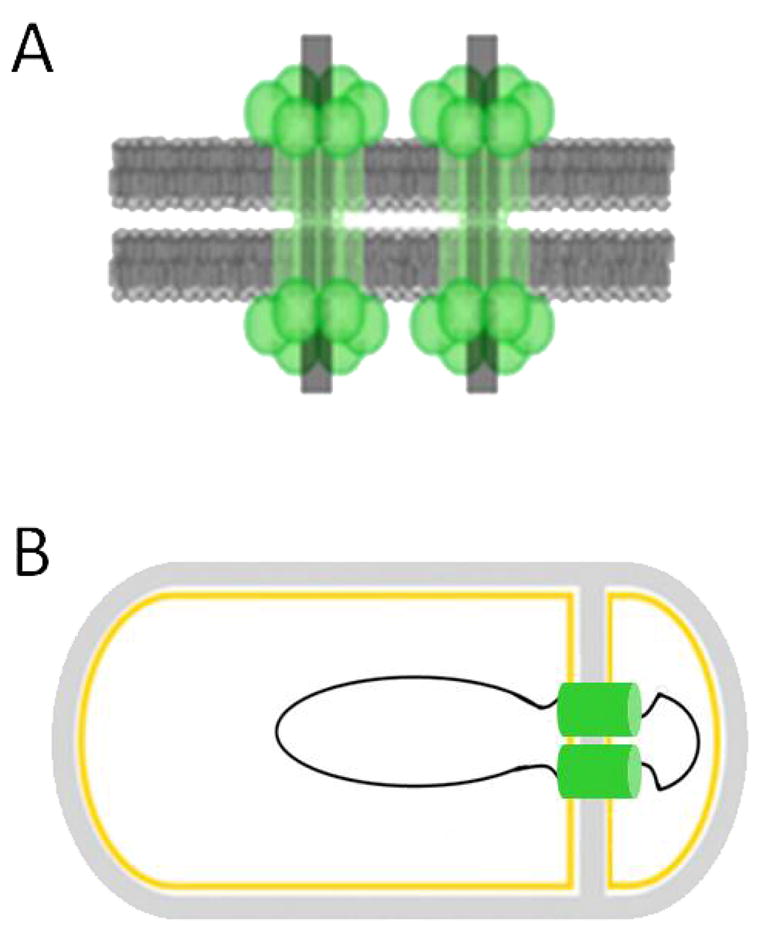
(A) The origin proximal one-third of the chromosome is contained within the forespore immediately following asymmetric septation. The polytopic membrane protein SpoIIIE is necessary for the translocation of the remaining two-thirds of the chromosome into the forespore. (B) SpoIIIE may be organized as a double-barreled DNA translocase, with each pore containing a single arm of the chromosome. Figures adapted from (Becker & Pogliano, 2007, Burton, et al., 2007).
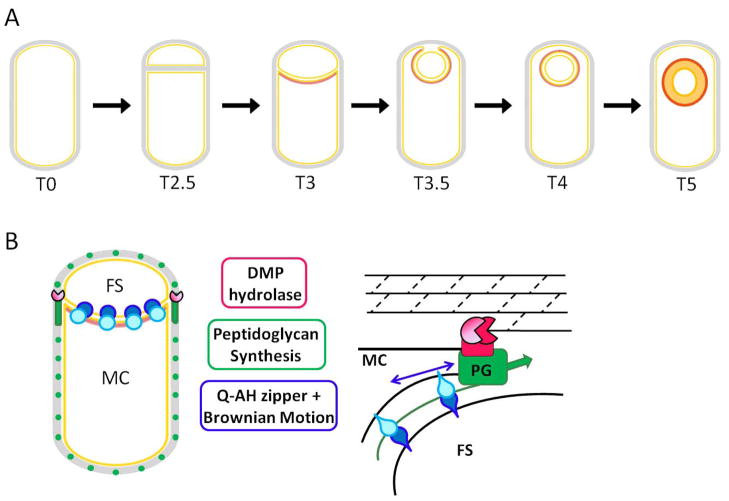
(A) Following asymmetric septation, the polar septum begins to curve and bend, resulting in its surrounding the forespore. The forespore detaches from the mother cell at the last step of engulfment (T4), which is immediately followed by spore cortex (thick gray band) synthesis. Under each stage is the approximate time in hours since the onset of sporulation at 37oC. (B) Three partially redundant molecular mechanisms are responsible for the membrane movement that occurs during sporulation: (1) The ‘DMP’ machine (red) is responsible for membrane migration by hydrolyzing the peptidoglycan between the two membranes (black lines), thereby moving the mother-cell derived outer forespore membrane around the forespore; (2) the SpoIIQ-SpoIIIAH zipper (blue) may function as a ratchet to irreversibly drive engulfment; and (3) peptidoglycan synthesis (green) of the germ cell wall could provide a motive force for membrane movement during early engulfment as well as the final step of forespore detachment. Figure adapted from (Meyer, et al., 2010).
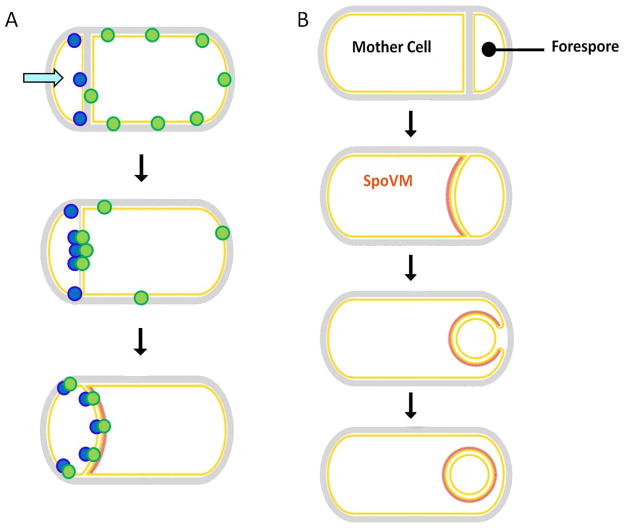
(A) The mother cell expressed protein SpoIIIAH (green) is initially present in the mother cell membrane but becomes progressively enriched at the sporulation septum through an interaction with the forespore expressed SpoIIQ protein (blue) that occurs in the space between the inner and outer sporulation membranes. Figure adapted from (Blaylock, et al., 2004). (B) The SpoVM protein is targeted to the outer forespore membrane because of its high negative curvature. Figure adapted from (Ramamurthi, 2010).
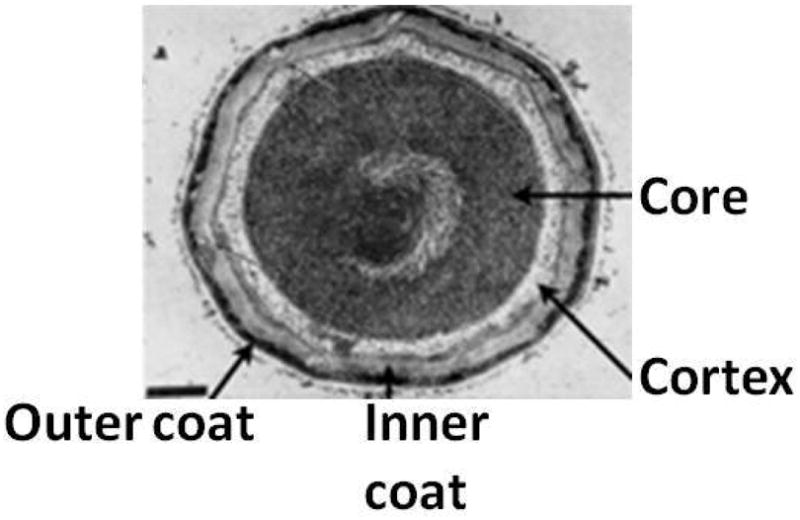
Multiple layers including the spore cortex composed of peptidoglycan and the inner and outer spore coats surround the spore core containing the compacted chromosome. Figure adapted from (Santo & Doi, 1974).
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
