

CDKB2 is involved in mitosis and DNA damage response in rice
- PMID: 22092531
- PMCID: PMC3440594
- DOI: 10.1111/j.1365-313X.2011.04847.x
CDKB2 is involved in mitosis and DNA damage response in rice
Abstract
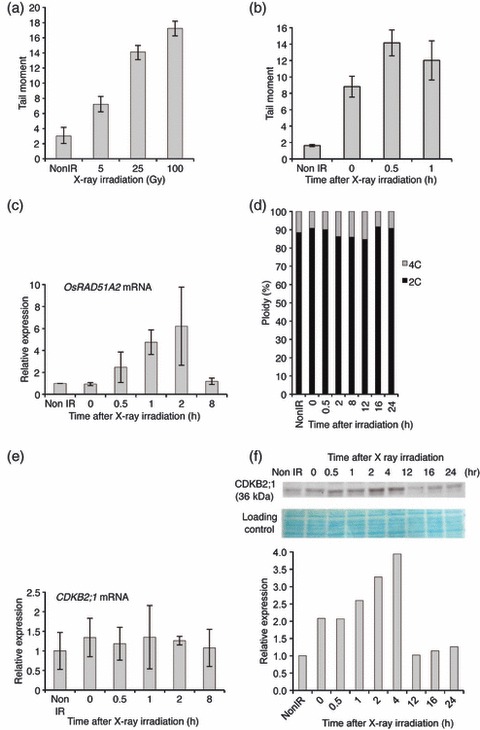
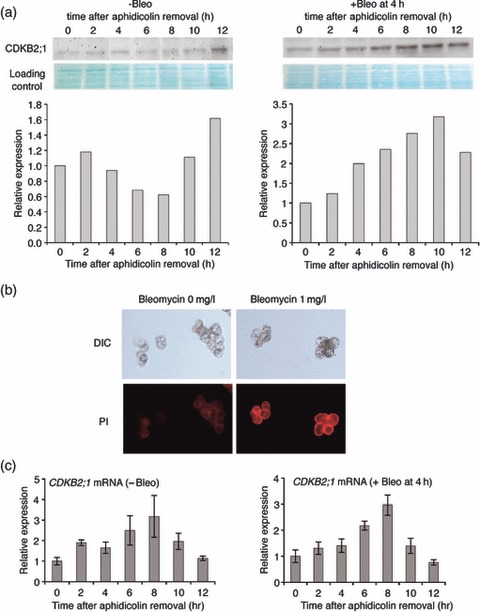
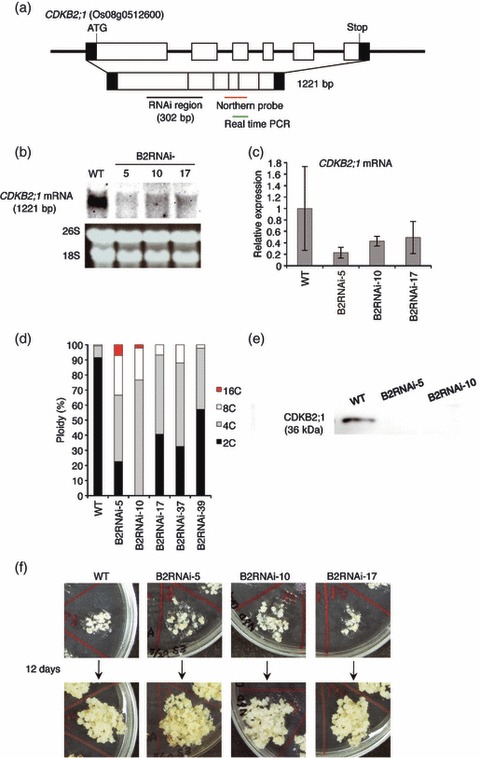
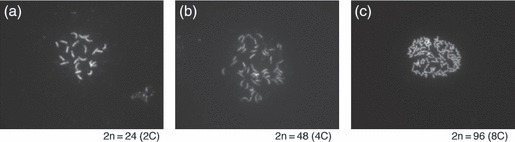
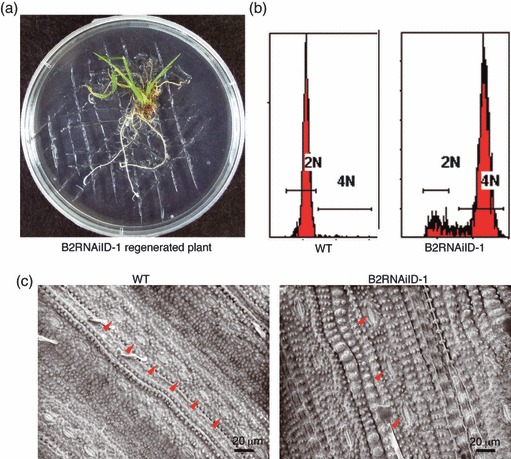
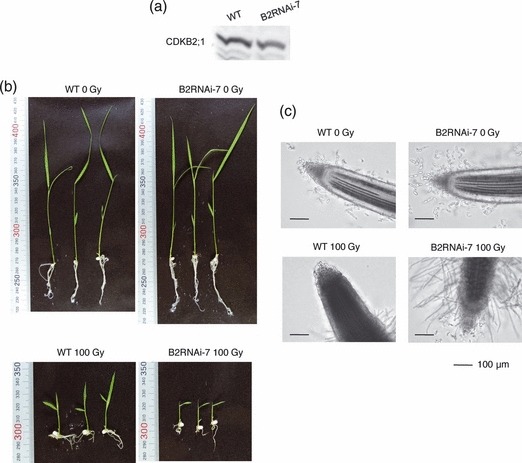
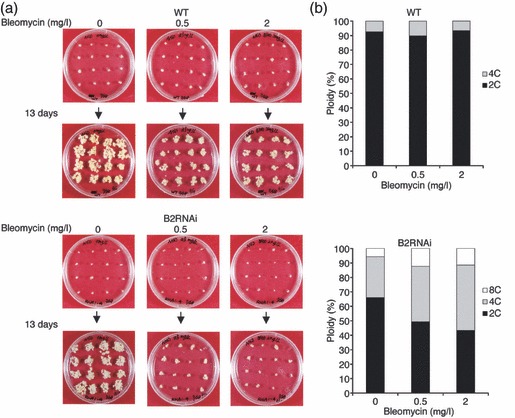
DNA damage checkpoints delay mitotic cell-cycle progression in response to DNA stress, stalling the cell cycle to allow time for repair. CDKB is a plant-specific cyclin-dependent kinase (CDK) that is required for the G₂/M transition of the cell cycle. In Arabidopsis, DNA damage leads the degradation of CDKB2, and the subsequent G₂ arrest gives cells time to repair damaged DNA. G₂ arrest also triggers transition from the mitotic cycle to endoreduplication, leading to the presence of polyploid cells in many tissues. In contrast, in rice (Oryza sativa), polyploid cells are found only in the endosperm. It was unclear whether endoreduplication contributes to alleviating DNA damage in rice (Oryza sativa). Here, we show that DNA damage neither down-regulates Orysa;CDKB2;1 nor induces endoreduplication in rice. Furthermore, we found increased levels of Orysa;CDKB2;1 protein upon DNA damage. These results suggest that CDKB2 functions differently in Arabidopsis and rice in response to DNA damage. Arabidopsis may adopt endoreduplication as a survival strategy under genotoxic stress conditions, but rice may enhance DNA repair capacity upon genotoxic stress. In addition, polyploid cells due to endomitosis were present in CDKB2;1 knockdown rice, suggesting an important role for Orysa;CDKB2;1 during mitosis.
© 2011 The Authors. The Plant Journal © 2011 Blackwell Publishing Ltd.
Figures
References
-
- Abe M, Kuroshita H, Umeda M, Itoh J, Nagato Y. The rice FLATTENED SHOOT MERISTEM, encoding CAF–1 p150 subunit, is required for meristem maintenance by regulating the cell-cycle period. Dev. Biol. 2008;319:384–393. - PubMed
-
- Baba A, Hasezawa S, Syono K. Cultivation of rice protoplasts and their transformation mediated by Agrobacterium spheroplasts. Plant Cell Physiol. 1986;27:463–471.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
