

MicroRNA transgene overexpression complements deficiency-based modifier screens in Drosophila
- PMID: 22095085
- PMCID: PMC3276628
- DOI: 10.1534/genetics.111.136689
MicroRNA transgene overexpression complements deficiency-based modifier screens in Drosophila
Abstract
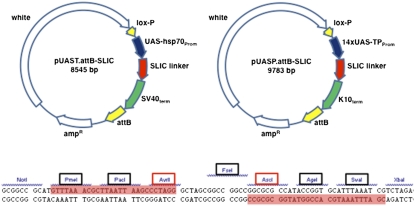
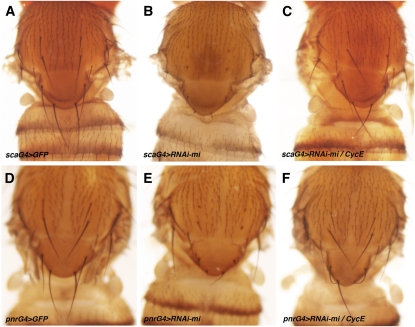
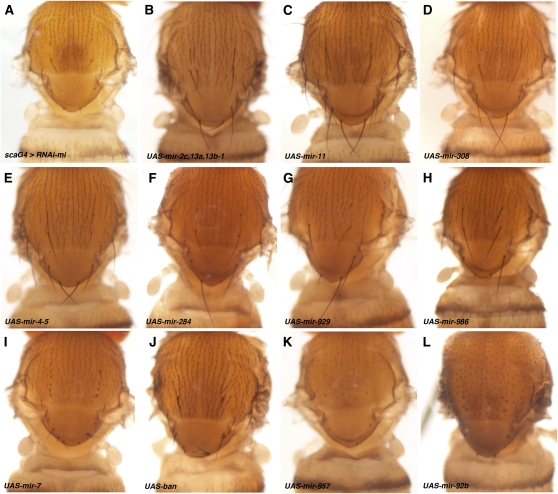
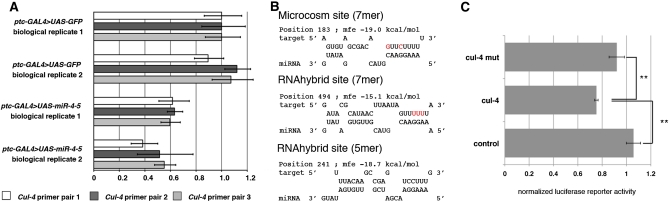
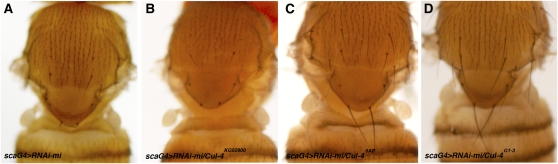
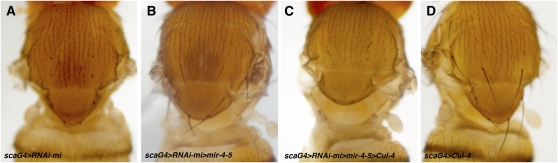
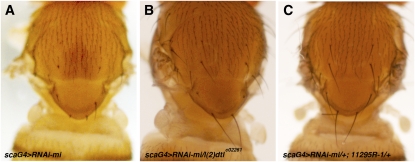
Dosage-sensitive modifier screening is a powerful tool for linking genes to biological processes. Use of chromosomal deletions permits sampling the effects of removing groups of genes related by position on the chromosome. Here, we explore the use of inducible microRNA transgenes as a complement to deficiency-based modifier screens. miRNAs are predicted to have hundreds of targets. miRNA overexpression provides an efficient means to reduces expression of large gene sets. A collection of transgenes was prepared to allow overexpression of 89 miRNAs or miRNA clusters. These transgenes and a set of genomic deficiencies were screened for their ability to modify the bristle phenotype of the cell-cycle regulator minus. Sixteen miRNAs were identified as dominant suppressors, while the deficiency screen uncovered four genomic regions that contain a dominant suppressor. Comparing the genes uncovered by the deletions with predicted miRNA targets uncovered a small set of candidate suppressors. Two candidates were identified as suppressors of the minus phenotype, Cullin-4 and CG5199/Cut8. Additionally, we show that Cullin-4 acts through its substrate receptor Cdt2 to suppress the minus phenotype. We suggest that inducible microRNA transgenes are a useful complement to deficiency-based modifier screens.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
