

Regulation of transforming growth factor-β1-driven lung fibrosis by galectin-3
- PMID: 22095546
- PMCID: PMC3410728
- DOI: 10.1164/rccm.201106-0965OC
Regulation of transforming growth factor-β1-driven lung fibrosis by galectin-3
Abstract
Rationale: Idiopathic pulmonary fibrosis (IPF) is a chronic dysregulated response to alveolar epithelial injury with differentiation of epithelial cells and fibroblasts into matrix-secreting myofibroblasts resulting in lung scaring. The prognosis is poor and there are no effective therapies or reliable biomarkers. Galectin-3 is a β-galactoside binding lectin that is highly expressed in fibrotic tissue of diverse etiologies.
Objectives: To examine the role of galectin-3 in pulmonary fibrosis.
Methods: We used genetic deletion and pharmacologic inhibition in well-characterized murine models of lung fibrosis. Further mechanistic studies were performed in vitro and on samples from patients with IPF.
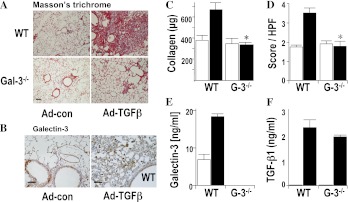
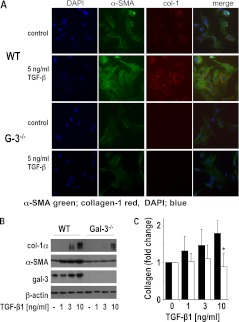
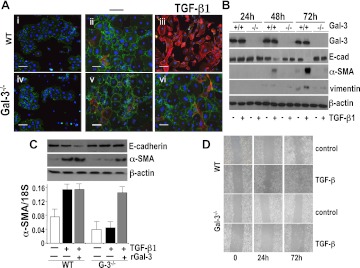
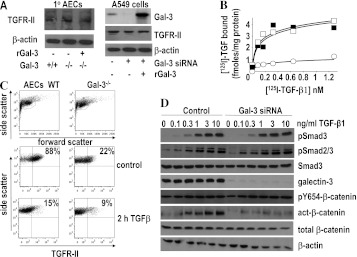
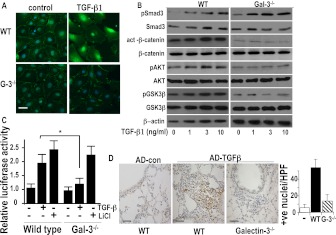
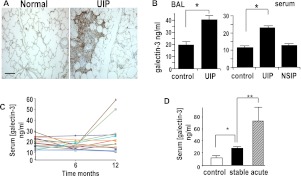
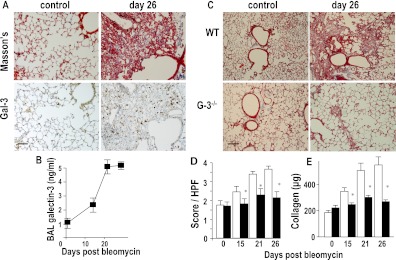
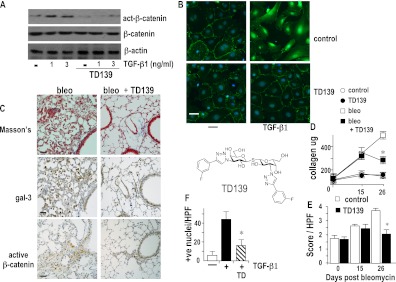
Measurements and main results: Transforming growth factor (TGF)-β and bleomycin-induced lung fibrosis was dramatically reduced in mice deficient in galectin-3, manifest by reduced TGF-β1-induced EMT and myofibroblast activation and collagen production. Galectin-3 reduced phosphorylation and nuclear translocation of β-catenin but had no effect on Smad2/3 phosphorylation. A novel inhibitor of galectin-3, TD139, blocked TGF-β-induced β-catenin activation in vitro and in vivo and attenuated the late-stage progression of lung fibrosis after bleomycin. There was increased expression of galectin-3 in the bronchoalveolar lavage fluid and serum from patients with stable IPF compared with nonspecific interstitial pneumonitis and controls, which rose sharply during an acute exacerbation suggesting that galectin-3 may be a marker of active fibrosis in IPF and that strategies that block galectin-3 may be effective in treating acute fibrotic exacerbations of IPF.
Conclusions: This study identifies galectin-3 as an important regulator of lung fibrosis and provides a proof of principle for galectin-3 inhibition as a potential novel therapeutic strategy for IPF.
Figures
Comment in
-
Galectin-3: the bridge over troubled waters.Am J Respir Crit Care Med. 2012 Mar 1;185(5):473-5. doi: 10.1164/rccm.201112-2190ED. Am J Respir Crit Care Med. 2012. PMID: 22383566 No abstract available.
References
-
- Gross TJ, Hunninghake GW. Idiopathic pulmonary fibrosis. N Engl J Med 2001;345:517–525 - PubMed
-
- Raghu G. Idiopathic pulmonary fibrosis: treatment options in pursuit of evidence-based approaches. Eur Respir J 2006;28:463–465 - PubMed
-
- Raghu G, Weycker D, Edelsberg J, Bradford WZ, Oster G. Incidence and prevalence of idiopathic pulmonary fibrosis. Am J Respir Crit Care Med 2006;174:810–816 - PubMed
-
- Selman M, Ruiz V, Cabrera S, Segura L, Ramirez R, Barrios R, Pardo A. TIMP-1, -2, -3, and -4 in idiopathic pulmonary fibrosis. A prevailing nondegradative lung microenvironment? Am J Physiol Lung Cell Mol Physiol 2000;279:L562–L574 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
