

Differential plasticity of epiblast and primitive endoderm precursors within the ICM of the early mouse embryo
- PMID: 22096072
- PMCID: PMC3231774
- DOI: 10.1242/dev.067702
Differential plasticity of epiblast and primitive endoderm precursors within the ICM of the early mouse embryo
Abstract
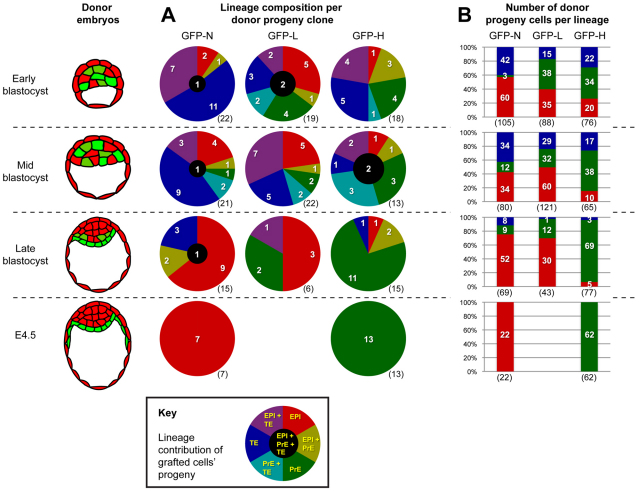
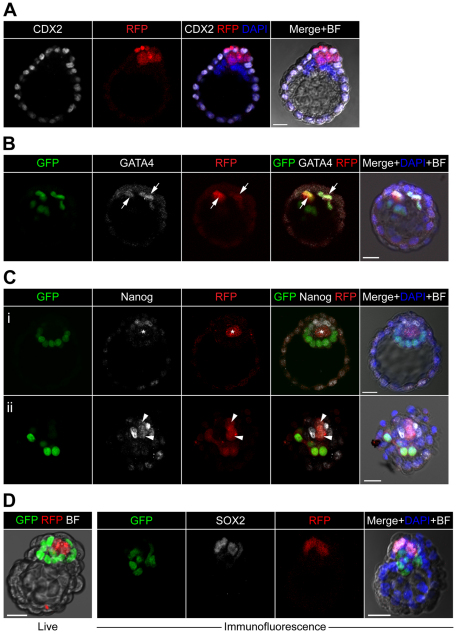
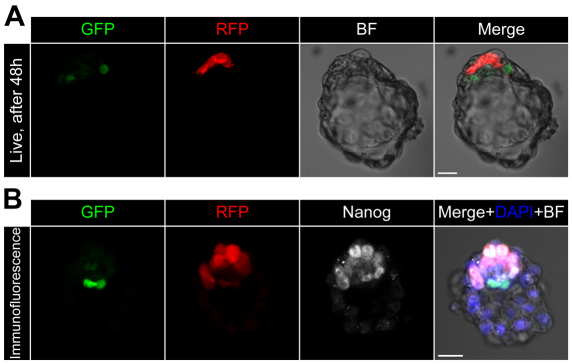
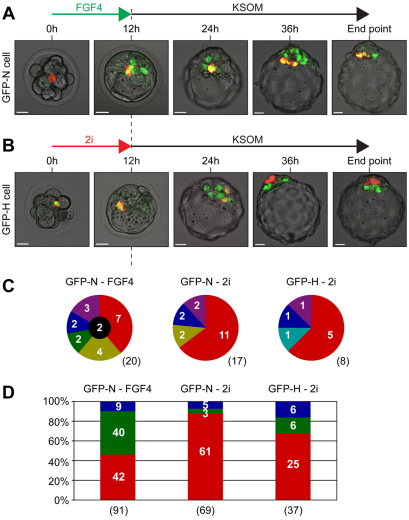
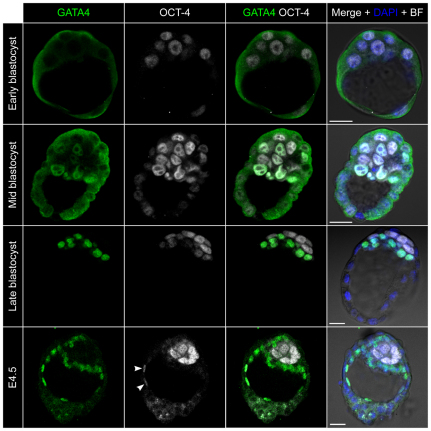
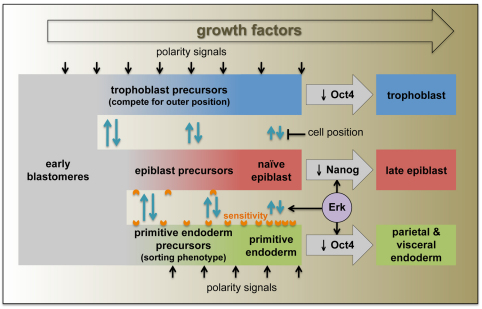
Cell differentiation during pre-implantation mammalian development involves the formation of two extra-embryonic lineages: trophoblast and primitive endoderm (PrE). A subset of cells within the inner cell mass (ICM) of the blastocyst does not respond to differentiation signals and forms the pluripotent epiblast, which gives rise to all of the tissues in the adult body. How this group of cells is set aside remains unknown. Recent studies documented distinct sequential phases of marker expression during the segregation of epiblast and PrE within the ICM. However, the connection between marker expression and lineage commitment remains unclear. Using a fluorescent reporter for PrE, we investigated the plasticity of epiblast and PrE precursors. Our observations reveal that loss of plasticity does not coincide directly with lineage restriction of epiblast and PrE markers, but rather with exclusion of the pluripotency marker Oct4 from the PrE. We note that individual ICM cells can contribute to all three lineages of the blastocyst until peri-implantation. However, epiblast precursors exhibit less plasticity than precursors of PrE, probably owing to differences in responsiveness to extracellular signalling. We therefore propose that the early embryo environment restricts the fate choice of epiblast but not PrE precursors, thus ensuring the formation and preservation of the pluripotent foetal lineage.
Figures

References
-
- Brons I. G., Smithers L. E., Trotter M. W., Rugg-Gunn P., Sun B., Chuva de Sousa Lopes S. M., Howlett S. K., Clarkson A., Ahrlund-Richter L., Pedersen R. A., et al. (2007). Derivation of pluripotent epiblast stem cells from mammalian embryos. Nature 448, 191–185 - PubMed
-
- Chambers I., Colby D., Robertson M., Nichols J., Lee S., Tweedie S., Smith A. (2003). Functional expression cloning of Nanog, a pluripotency sustaining factor in embryonic stem cells. Cell 113, 643–655 - PubMed
-
- Chazaud C., Yamanaka Y., Pawson T., Rossant J. (2006). Early lineage segregation between epiblast and primitive endoderm in mouse blastocysts through the Grb2-MAPK pathway. Dev. Cell 10, 615–624 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 079249/WT_/Wellcome Trust/United Kingdom
- BP R107861/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- G0800784/MRC_/Medical Research Council/United Kingdom
- BB/G012393/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- R01-DK084391/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
