

Integrin-αvβ3 regulates thrombopoietin-mediated maintenance of hematopoietic stem cells
- PMID: 22096247
- PMCID: PMC3251239
- DOI: 10.1182/blood-2011-02-335430
Integrin-αvβ3 regulates thrombopoietin-mediated maintenance of hematopoietic stem cells
Abstract
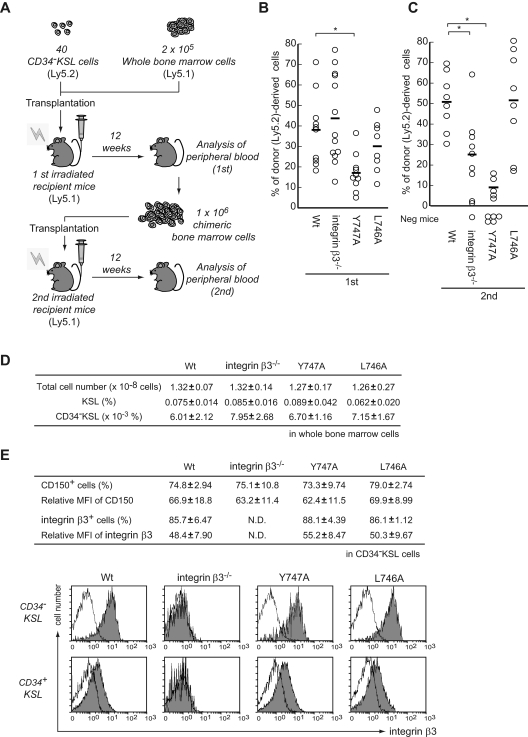
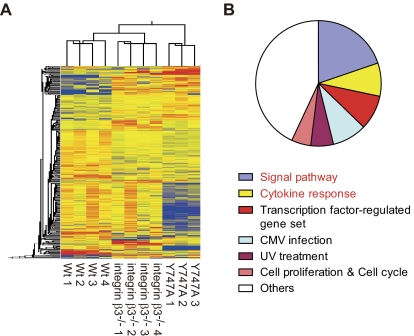
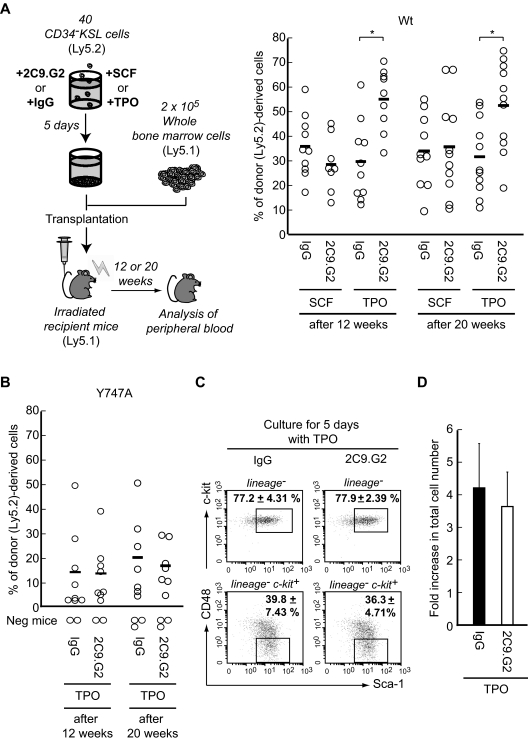
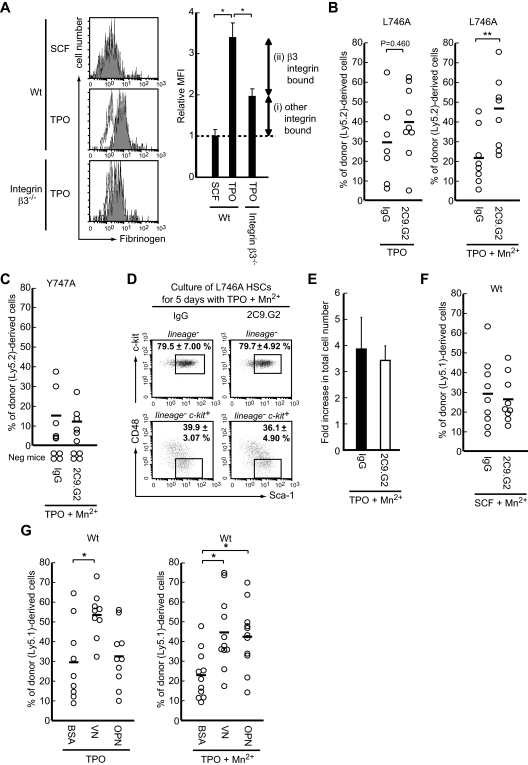
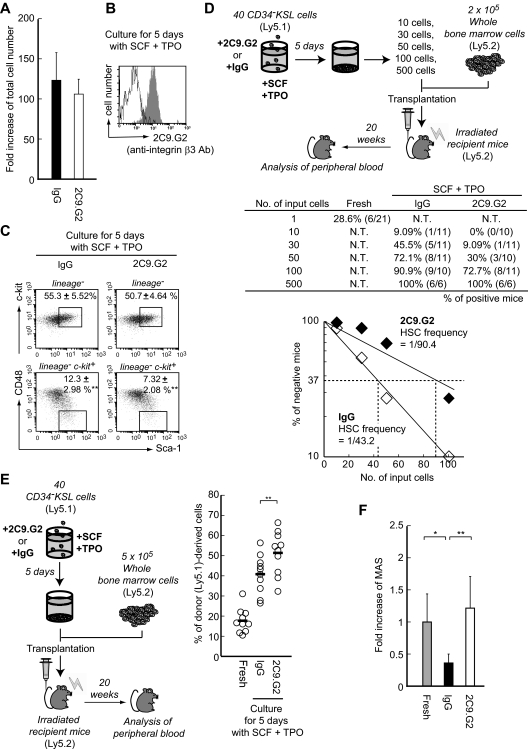
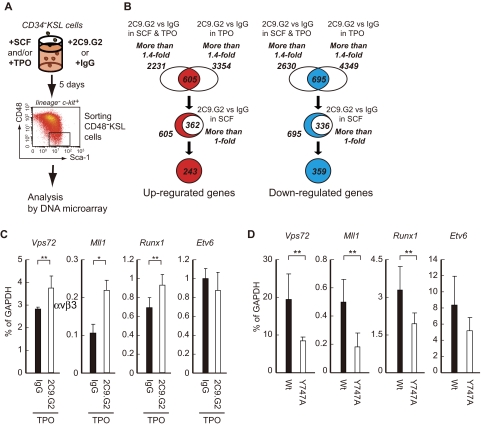
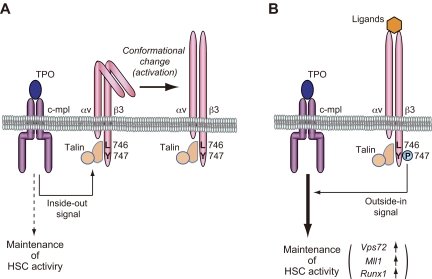
Throughout life, one's blood supply depends on sustained division of hematopoietic stem cells (HSCs) for self-renewal and differentiation. Within the bone marrow microenvironment, an adhesion-dependent or -independent niche system regulates HSC function. Here we show that a novel adhesion-dependent mechanism via integrin-β3 signaling contributes to HSC maintenance. Specific ligation of β3-integrin on HSCs using an antibody or extracellular matrix protein prevented loss of long-term repopulating (LTR) activity during ex vivo culture. The actions required activation of αvβ3-integrin "inside-out" signaling, which is dependent on thrombopoietin (TPO), an essential cytokine for activation of dormant HSCs. Subsequent "outside-in" signaling via phosphorylation of Tyr747 in the β3-subunit cytoplasmic domain was indispensable for TPO-dependent, but not stem cell factor-dependent, LTR activity in HSCs in vivo. This was accompanied with enhanced expression of Vps72, Mll1, and Runx1, 3 factors known to be critical for maintaining HSC activity. Thus, our findings demonstrate a mechanistic link between β3-integrin and TPO in HSCs, which may contribute to maintenance of LTR activity in vivo as well as during ex vivo culture.
Figures
Similar articles
-
Nov/CCN3 regulates long-term repopulating activity of murine hematopoietic stem cells via integrin αvβ3.Int J Hematol. 2014 Apr;99(4):393-406. doi: 10.1007/s12185-014-1534-x. Epub 2014 Feb 22. Int J Hematol. 2014. PMID: 24563081 Free PMC article.
-
Integrin αvβ3 enhances the suppressive effect of interferon-γ on hematopoietic stem cells.EMBO J. 2017 Aug 15;36(16):2390-2403. doi: 10.15252/embj.201796771. Epub 2017 Jul 3. EMBO J. 2017. PMID: 28673932 Free PMC article.
-
Lnk negatively regulates self-renewal of hematopoietic stem cells by modifying thrombopoietin-mediated signal transduction.Proc Natl Acad Sci U S A. 2007 Feb 13;104(7):2349-54. doi: 10.1073/pnas.0606238104. Epub 2007 Feb 6. Proc Natl Acad Sci U S A. 2007. PMID: 17284614 Free PMC article.
-
Thrombopoietin and hematopoietic stem cells.Cell Cycle. 2011 May 15;10(10):1582-9. doi: 10.4161/cc.10.10.15619. Epub 2011 May 15. Cell Cycle. 2011. PMID: 21478671 Free PMC article. Review.
-
Quantitative assessment of the stem cell self-renewal capacity.Ann N Y Acad Sci. 2001 Jun;938:18-24; discussion 24-5. doi: 10.1111/j.1749-6632.2001.tb03570.x. Ann N Y Acad Sci. 2001. PMID: 11458506 Review.
Cited by
-
Non-Triple Helical Form of Type IV Collagen α1 Chain.Heliyon. 2015 Dec 9;1(4):e00051. doi: 10.1016/j.heliyon.2015.e00051. eCollection 2015 Dec. Heliyon. 2015. PMID: 27441234 Free PMC article.
-
Physiological Cues Involved in the Regulation of Adhesion Mechanisms in Hematopoietic Stem Cell Fate Decision.Front Cell Dev Biol. 2020 Jul 10;8:611. doi: 10.3389/fcell.2020.00611. eCollection 2020. Front Cell Dev Biol. 2020. PMID: 32754597 Free PMC article. Review.
-
Haematopoietic stem cell self-renewal in vivo and ex vivo.Nat Rev Genet. 2020 Sep;21(9):541-554. doi: 10.1038/s41576-020-0241-0. Epub 2020 May 28. Nat Rev Genet. 2020. PMID: 32467607 Free PMC article. Review.
-
Secreted protein Del-1 regulates myelopoiesis in the hematopoietic stem cell niche.J Clin Invest. 2017 Oct 2;127(10):3624-3639. doi: 10.1172/JCI92571. Epub 2017 Aug 28. J Clin Invest. 2017. PMID: 28846069 Free PMC article.
-
Thrombopoietin protects hematopoietic stem cells from retrotransposon-mediated damage by promoting an antiviral response.J Exp Med. 2018 May 7;215(5):1463-1480. doi: 10.1084/jem.20170997. Epub 2018 Apr 3. J Exp Med. 2018. PMID: 29615469 Free PMC article.
References
-
- Arai F, Hirao A, Ohmura M, et al. Tie2/angiopoietin-1 signaling regulates hematopoietic stem cell quiescence in the bone marrow niche. Cell. 2004;118(2):149–161. - PubMed
-
- Nilsson SK, Johnston HM, Whitty GA, et al. Osteopontin, a key component of the hematopoietic stem cell niche and regulator of primitive hematopoietic progenitor cells. Blood. 2005;106(4):1232–1239. - PubMed
-
- Qian H, Buza-Vidas N, Hyland CD, et al. Critical role of thrombopoietin in maintaining adult quiescent hematopoietic stem cells. Cell Stem Cell. 2007;1(6):671–684. - PubMed
-
- Yoshihara H, Arai F, Hosokawa K, et al. Thrombopoietin/MPL signaling regulates hematopoietic stem cell quiescence and interaction with the osteoblastic niche. Cell Stem Cell. 2007;1(6):685–697. - PubMed
-
- Yamazaki S, Iwama A, Takayanagi S, Eto K, Ema H, Nakauchi H. TGF-beta as a candidate bone marrow niche signal to induce hematopoietic stem cell hibernation. Blood. 2009;113(6):1250–1256. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
